
Respiratory system facts for kids
Quick facts for kids Respiratory system |
|
|---|---|
 |
|
| A complete, schematic view of the human respiratory system with their parts and functions. | |
| Latin | systema respiratorium |
- For the biochemical process, see respiration
The respiratory system, also called the gas exchange system, is the body getting rid of carbon dioxide and taking in oxygen. Carbon dioxide, a waste product, goes out of the body. Oxygen, which the body needs, comes in.
The first step in this process is breathing in air, or inhaling. The taking in of air rich in oxygen into the body is called inhalation and giving out of air rich in carbon dioxide from the body is called exhalation. The second step is gas exchange in the lungs where oxygen is diffused into the blood and the carbon dioxide diffuses out of the blood. The third process is cellular respiration, which produces the chemical energy that the cells in the body need, and carbon dioxide. Finally, the carbon dioxide from cellular respiration is breathed out of body from the lungs.
In most fish, and a number of other aquatic animals (both vertebrates and invertebrates), the respiratory system consists of gills, which are either partially or completely external organs, bathed in the watery environment. This water flows over the gills by a variety of active or passive means. Gas exchange takes place in the gills which consist of thin or very flat filaments and lammellae which expose a very large surface area of highly vascularized tissue to the water.
Other animals, such as insects, have respiratory systems with very simple anatomical features, and in amphibians, even the skin plays a vital role in gas exchange. Plants also have respiratory systems but the directionality of gas exchange can be opposite to that in animals. The respiratory system in plants includes anatomical features such as stomata, that are found in various parts of the plant.
Contents
Breathing

For respiration to happen, the body needs a constant supply of oxygen, which is done by breathing. Inhalation is the breathing in of air. To inhale, the lungs expand, decreasing the air pressure in the lungs. This is caused by the diaphragm (a sheet of muscular tissue that separates the lungs from the abdomen) and the muscles between the ribs contracting to expand the chest, which also expands the lungs. As the air pressure inside the lungs are lower when it has expanded, air from outside at higher pressure comes rushing into the area of low pressure in the lungs. Air first passes through the nose and mouth, then through the larynx (voice box), then down the trachea (windpipe), and into the lungs and comes out
The lungs are made of many tubes or branches. As air enters the lungs, it first goes through branches called the bronchi, then through smaller branches called bronchioles, and finally into the air sacs. Gas exchange occurs in the air sacs where oxygen is exchanged with carbon dioxide. The carbon dioxide in the air sacs now need to be exhaled, or breathed out. In the reverse process to inhaling, the diaphragm and the rib muscles relax, causing the lungs to be smaller. As the air pressure in the lungs is greater when the lungs are smaller, air is forced out. The exhaled air has a high concentration of carbon dioxide and a low concentration of oxygen. The maximum volume of air that can be inhaled and exhaled is called the vital capacity of the lungs and is up to five liters.
Gas exchange

The inhaled air goes down to the air sacs at the end of each bronchiole. The air sacs are called alveoli — they have a large surface area, and are moist, thin, and close to a blood supply. The inhaled air has a much greater concentration of oxygen than carbon dioxide whilst the blood flowing to the lungs has a more carbon dioxide than oxygen. This creates a concentration gradient between the air in the air sacs and the blood, meaning there is more oxygen in the air than the blood. As the membrane, oxygen can easily diffuse in and out. Oxygen at high concentration in the air sacs diffuses into the blood where oxygen concentration is low, and carbon dioxide at high concentration in the blood diffuses into the air sacs where carbon dioxide concentration is low. The oxygen in the blood enters the circulatory system and is used by the cells in the body. The carbon dioxide in the air sacs are exhaled out of the body.
Exceptional mammals
Horses
Horses are obligate nasal breathers which means that they are different from many other mammals because they do not have the option of breathing through their mouths and must take in air through their noses. A flap of tissue called the soft palate blocks off the pharynx from the mouth (oral cavity) of the horse, except when swallowing. This helps to prevent the horse from inhaling food, but does not allow use of the mouth to breathe when in respiratory distress, a horse can only breathe through its nostrils.
Elephants
The elephant is the only mammal known to have no pleural space. Instead, the parietal and visceral pleura are both composed of dense connective tissue and joined to each other via loose connective tissue. This lack of a pleural space, along with an unusually thick diaphragm, are thought to be evolutionary adaptations allowing the elephant to remain underwater for long periods while breathing through its trunk which emerges as a snorkel.
In the elephant the lungs are attached to the diaphragm and breathing relies mainly on the diaphragm rather than the expansion of the ribcage.
Birds



Key:
1. skull; 2. cervical vertebrae; 3. furcula; 4. coracoid; 5. vertebral ribs; 6. sternum and its keel; 7. patella; 8. tarsus; 9. digits; 10. tibia (tibiotarsus); 11. fibula (tibiotarsus); 12. femur; 13. ischium (innominate); 14. pubis (innominate); 15. ilium (innominate); 16. caudal vertebrae; 17. pygostyle; 18. synsacrum; 19. scapula; 20. dorsal vertebrae; 21. humerus; 22. ulna; 23. radius; 24. carpus (carpometacarpus); 25. metacarpus (carpometacarpus); 26. digits; 27. alula
The respiratory system of birds differs significantly from that found in mammals. Firstly, they have rigid lungs which do not expand and contract during the breathing cycle. Instead an extensive system of air sacs (Fig. 15) distributed throughout their bodies act as the bellows drawing environmental air into the sacs, and expelling the spent air after it has passed through the lungs (Fig. 18). Birds also do not have diaphragms or pleural cavities.
Bird lungs are smaller than those in mammals of comparable size, but the air sacs account for 15% of the total body volume, compared to the 7% devoted to the alveoli which act as the bellows in mammals.
Inhalation and exhalation are brought about by alternately increasing and decreasing the volume of the entire thoraco-abdominal cavity (or coelom) using both their abdominal and costal muscles. During inhalation the muscles attached to the vertebral ribs (Fig. 17) contract angling them forwards and outwards. This pushes the sternal ribs, to which they are attached at almost right angles, downwards and forwards, taking the sternum (with its prominent keel) in the same direction (Fig. 17). This increases both the vertical and transverse diameters of thoracic portion of the trunk. The forward and downward movement of, particularly, the posterior end of the sternum pulls the abdominal wall downwards, increasing the volume of that region of the trunk as well. The increase in volume of the entire trunk cavity reduces the air pressure in all the thoraco-abdominal air sacs, causing them to fill with air as described below.
During exhalation the external oblique muscle which is attached to the sternum and vertebral ribs anteriorly, and to the pelvis (pubis and ilium in Fig. 17) posteriorly (forming part of the abdominal wall) reverses the inhalatory movement, while compressing the abdominal contents, thus increasing the pressure in all the air sacs. Air is therefore expelled from the respiratory system in the act of exhalation.

During inhalation air enters the trachea via the nostrils and mouth, and continues to just beyond the syrinx at which point the trachea branches into two primary bronchi, going to the two lungs (Fig. 16). The primary bronchi enter the lungs to become the intrapulmonary bronchi, which give off a set of parallel branches called ventrobronchi and, a little further on, an equivalent set of dorsobronchi (Fig. 16). The ends of the intrapulmonary bronchi discharge air into the posterior air sacs at the caudal end of the bird. Each pair of dorso-ventrobronchi is connected by a large number of parallel microscopic air capillaries (or parabronchi) where gas exchange occurs (Fig. 16). As the bird inhales, tracheal air flows through the intrapulmonary bronchi into the posterior air sacs, as well as into the dorsobronchi, but not into the ventrobronchi (Fig. 18). This is due to the bronchial architecture which directs the inhaled air away from the openings of the ventrobronchi, into the continuation of the intrapulmonary bronchus towards the dorsobronchi and posterior air sacs. From the dorsobronchi the inhaled air flows through the parabronchi (and therefore the gas exchanger) to the ventrobronchi from where the air can only escape into the expanding anterior air sacs. So, during inhalation, both the posterior and anterior air sacs expand, the posterior air sacs filling with fresh inhaled air, while the anterior air sacs fill with "spent" (oxygen-poor) air that has just passed through the lungs.

During exhalation the pressure in the posterior air sacs (which were filled with fresh air during inhalation) increases due to the contraction of the oblique muscle described above. The aerodynamics of the interconnecting openings from the posterior air sacs to the dorsobronchi and intrapulmonary bronchi ensures that the air leaves these sacs in the direction of the lungs (via the dorsobronchi), rather than returning down the intrapulmonary bronchi (Fig. 18). From the dorsobronchi the fresh air from the posterior air sacs flows through the parabronchi (in the same direction as occurred during inhalation) into ventrobronchi. The air passages connecting the ventrobronchi and anterior air sacs to the intrapulmonary bronchi direct the "spent", oxygen poor air from these two organs to the trachea from where it escapes to the exterior. Oxygenated air therefore flows constantly (during the entire breathing cycle) in a single direction through the parabronchi.
The blood flow through the bird lung is at right angles to the flow of air through the parabronchi, forming a cross-current flow exchange system (Fig. 19). The partial pressure of oxygen in the parabronchi declines along their lengths as O2 diffuses into the blood. The blood capillaries leaving the exchanger near the entrance of airflow take up more O2 than do the capillaries leaving near the exit end of the parabronchi. When the contents of all capillaries mix, the final partial pressure of oxygen of the mixed pulmonary venous blood is higher than that of the exhaled air, but is nevertheless less than half that of the inhaled air, thus achieving roughly the same systemic arterial blood partial pressure of oxygen as mammals do with their bellows-type lungs.
The trachea is an area of dead space: the oxygen-poor air it contains at the end of exhalation is the first air to re-enter the posterior air sacs and lungs. In comparison to the mammalian respiratory tract, the dead space volume in a bird is, on average, 4.5 times greater than it is in mammals of the same size. Birds with long necks will inevitably have long tracheae, and must therefore take deeper breaths than mammals do to make allowances for their greater dead space volumes. In some birds (e.g. the whooper swan, Cygnus cygnus, the white spoonbill, Platalea leucorodia, the whooping crane, Grus americana, and the helmeted curassow, Pauxi pauxi) the trachea, which some cranes can be 1.5 m long, is coiled back and forth within the body, drastically increasing the dead space ventilation. The purpose of this extraordinary feature is unknown.
Reptiles
The anatomical structure of the lungs is less complex in reptiles than in mammals, with reptiles lacking the very extensive airway tree structure found in mammalian lungs. Gas exchange in reptiles still occurs in alveoli however. Reptiles do not possess a diaphragm. Thus, breathing occurs via a change in the volume of the body cavity which is controlled by contraction of intercostal muscles in all reptiles except turtles. In turtles, contraction of specific pairs of flank muscles governs inhalation and exhalation.
Amphibians
Both the lungs and the skin serve as respiratory organs in amphibians. The ventilation of the lungs in amphibians relies on positive pressure ventilation. Muscles lower the floor of the oral cavity, enlarging it and drawing in air through the nostrils into the oral cavity. With the nostrils and mouth closed, the floor of the oral cavity is then pushed up, which forces air down the trachea into the lungs. The skin of these animals is highly vascularized and moist, with moisture maintained via secretion of mucus from specialised cells, and is involved in cutaneous respiration. While the lungs are of primary organs for gas exchange between the blood and the environmental air (when out of the water), the skin's unique properties aid rapid gas exchange when amphibians are submerged in oxygen-rich water. Some amphibians have gills, either in the early stages of their development (e.g. tadpoles of frogs), while others retain them into adulthood (e.g. some salamanders).
Fish
.jpg)


Oxygen is poorly soluble in water. Fully aerated fresh water therefore contains only 8–10 ml O2/liter compared to the O2 concentration of 210 ml/liter in the air at sea level. Furthermore, the coefficient of diffusion (i.e. the rate at which a substances diffuses from a region of high concentration to one of low concentration, under standard conditions) of the respiratory gases is typically 10,000 faster in air than in water. Thus oxygen, for instance, has a diffusion coefficient of 17.6 mm2/s in air, but only 0.0021 mm2/s in water. The corresponding values for carbon dioxide are 16 mm2/s in air and 0.0016 mm2/s in water. This means that when oxygen is taken up from the water in contact with a gas exchanger, it is replaced considerably more slowly by the oxygen from the oxygen-rich regions small distances away from the exchanger than would have occurred in air. Fish have developed gills deal with these problems. Gills are specialized organs containing filaments, which further divide into lamellae. The lamellae contain a dense thin walled capillary network that exposes a large gas exchange surface area to the very large volumes of water passing over them.
Gills use a countercurrent exchange system that increases the efficiency of oxygen-uptake from the water. Fresh oxygenated water taken in through the mouth is uninterruptedly "pumped" through the gills in one direction, while the blood in the lamellae flows in the opposite direction, creating the countercurrent blood and water flow (Fig. 22), on which the fish's survival depends.
Water is drawn in through the mouth by closing the operculum (gill cover), and enlarging the mouth cavity (Fig. 23). Simultaneously the gill chambers enlarge, producing a lower pressure there than in the mouth causing water to flow over the gills. The mouth cavity then contracts, inducing the closure of the passive oral valves, thereby preventing the back-flow of water from the mouth (Fig. 23). The water in the mouth is, instead, forced over the gills, while the gill chambers contract emptying the water they contain through the opercular openings (Fig. 23). Back-flow into the gill chamber during the inhalatory phase is prevented by a membrane along the ventroposterior border of the operculum (diagram on the left in Fig. 23). Thus the mouth cavity and gill chambers act alternately as suction pump and pressure pump to maintain a steady flow of water over the gills in one direction. Since the blood in the lamellar capillaries flows in the opposite direction to that of the water, the consequent countercurrent flow of blood and water maintains steep concentration gradients for oxygen and carbon dioxide along the entire length of each capillary (lower diagram in Fig. 22). Oxygen is, therefore, able to continually diffuse down its gradient into the blood, and the carbon dioxide down its gradient into the water. Although countercurrent exchange systems theoretically allow an almost complete transfer of a respiratory gas from one side of the exchanger to the other, in fish less than 80% of the oxygen in the water flowing over the gills is generally transferred to the blood.
In certain active pelagic sharks, water passes through the mouth and over the gills while they are moving, in a process known as "ram ventilation". While at rest, most sharks pump water over their gills, as most bony fish do, to ensure that oxygenated water continues to flow over their gills. But a small number of species have lost the ability to pump water through their gills and must swim without rest. These species are obligate ram ventilators and would presumably asphyxiate if unable to move. Obligate ram ventilation is also true of some pelagic bony fish species.
There are a few fish that can obtain oxygen for brief periods of time from air swallowed from above the surface of the water. Thus lungfish possess one or two lungs, and the labyrinth fish have developed a special "labyrinth organ", which characterizes this suborder of fish. The labyrinth organ is a much-folded suprabranchial accessory breathing organ. It is formed by a vascularized expansion of the epibranchial bone of the first gill arch, and is used for respiration in air. This organ allows labyrinth fish to take in oxygen directly from the air, instead of taking it from the water in which they reside through the use of gills. The labyrinth organ helps the oxygen in the inhaled air to be absorbed into the bloodstream. As a result, labyrinth fish can survive for a short period of time out of water, as they can inhale the air around them, provided they stay moist. Labyrinth fish are not born with functional labyrinth organs. The development of the organ is gradual and most juvenile labyrinth fish breathe entirely with their gills and develop the labyrinth organs when they grow older.
Invertebrates
Arthropods
Some species of crab use a respiratory organ called a branchiostegal lung. Its gill-like structure increases the surface area for gas exchange which is more suited to taking oxygen from the air than from water. Some of the smallest spiders and mites can breathe simply by exchanging gas through the surface of the body. Larger spiders, scorpions and other arthropods use a primitive book lung.
Insects
Most insects breath passively through their spiracles (special openings in the exoskeleton) and the air reaches every part of the body by means of a series of smaller and smaller tubes called 'trachaea' when their diameters are relatively large, and 'tracheoles' when their diameters are very small. The tracheoles make contact with individual cells throughout the body. They are partially filled with fluid, which can be withdrawn from the individual tracheoles when the tissues, such as muscles, are active and have a high demand for oxygen, bringing the air closer to the active cells. This is probably brought about by the buildup of lactic acid in the active muscles causing an osmotic gradient, moving the water out of the tracheoles and into the active cells. Diffusion of gases is effective over small distances but not over larger ones, this is one of the reasons insects are all relatively small. Insects which do not have spiracles and trachaea, such as some Collembola, breathe directly through their skins, also by diffusion of gases.
The number of spiracles an insect has is variable between species, however, they always come in pairs, one on each side of the body, and usually one pair per segment. Some of the Diplura have eleven, with four pairs on the thorax, but in most of the ancient forms of insects, such as Dragonflies and Grasshoppers there are two thoracic and eight abdominal spiracles. However, in most of the remaining insects, there are fewer. It is at the level of the tracheoles that oxygen is delivered to the cells for respiration.
Insects were once believed to exchange gases with the environment continuously by the simple diffusion of gases into the tracheal system. More recently, however, large variation in insect ventilatory patterns has been documented and insect respiration appears to be highly variable. Some small insects do not demonstrate continuous respiratory movements and may lack muscular control of the spiracles. Others, however, utilize muscular contraction of the abdomen along with coordinated spiracle contraction and relaxation to generate cyclical gas exchange patterns and to reduce water loss into the atmosphere. The most extreme form of these patterns is termed discontinuous gas exchange cycles.
Molluscs
Molluscs generally possess gills that allow gas exchange between the aqueous environment and their circulatory systems. These animals also possess a heart that pumps blood containing hemocyanin as its oxygen-capturing molecule. Hence, this respiratory system is similar to that of vertebrate fish. The respiratory system of gastropods can include either gills or a lung.
Plants
Plants use carbon dioxide gas in the process of photosynthesis, and exhale oxygen gas as waste. The chemical equation of photosynthesis is 6 CO2 (carbon dioxide) and 6 H2O (water), which in the presence of sunlight makes C6H12O6 (glucose) and 6 O2 (oxygen). Photosynthesis uses electrons on the carbon atoms as the repository for the energy obtained from sunlight. Respiration is the opposite of photosynthesis. It reclaims the energy to power chemical reactions in cells. In so doing the carbon atoms and their electrons are combined with oxygen forming CO2 which is easily removed from both the cells and the organism. Plants use both processes, photosynthesis to capture the energy and oxidative metabolism to use it.
Plant respiration is limited by the process of diffusion. Plants take in carbon dioxide through holes, known as stomata, that can open and close on the undersides of their leaves and sometimes other parts of their anatomy. Most plants require some oxygen for catabolic processes (break-down reactions that release energy). But the quantity of O2 used per hour is small as they are not involved in activities that require high rates of aerobic metabolism. Their requirement for air, however, is very high as they need CO2 for photosynthesis, which constitutes only 0.04% of the environmental air. Thus, to make 1 g of glucose requires the removal of all the CO2 from at least 18.7 liters of air at sea level. But inefficiencies in the photosynthetic process cause considerably greater volumes of air to be used.
Related pages
Images for kids
-
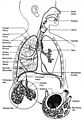
Fig. 1. Respiratory system
-
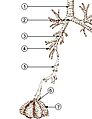 Fig. 2. The lower respiratory tract, or "Respiratory Tree"
Fig. 2. The lower respiratory tract, or "Respiratory Tree"- Trachea
- Mainstem bronchus
- Lobar bronchus
- Segmental bronchus
- Bronchiole
- Alveolar duct
- Alveolus
-

Fig. 14 A graph showing the relationship between total atmospheric pressure and altitude above sea level.
-

Fig. 13 Aerial photo of Mount Everest from the south, behind Nuptse and Lhotse.




See also
 In Spanish: Aparato respiratorio para niños
In Spanish: Aparato respiratorio para niños
