
Giardia duodenalis facts for kids
Quick facts for kids Giardia duodenalis |
|
|---|---|
 |
|
| Giardia lamblia cell, SEM | |
| Scientific classification |
|
| Domain: | Eukaryota |
| Phylum: | Metamonada |
| Order: | Diplomonadida |
| Family: | Hexamitidae |
| Genus: | Giardia |
| Species: |
G. duodenalis
|
| Binomial name | |
| Giardia duodenalis Stiles, 1902
|
|
| Script error: The function "autoWithCaption" does not exist. | |
| Synonyms | |
|
|
Script error: No such module "Check for conflicting parameters".
Giardia duodenalis, also called Giardia intestinalis or Giardia lamblia, is a tiny parasite. It has whip-like tails called flagella. This parasite lives and grows in the small intestine of humans and animals. It causes a tummy illness called giardiasis.
The parasite sticks to the inside of the intestine. It uses a special sticky disc. Giardia makes more of itself by splitting in two. This is called binary fission. It stays only in the small intestine. It does not spread through the blood or to other body parts.
Giardia has a strong outer layer. This helps it survive outside a body. It can even survive some cleaning chemicals like chlorine. Giardia gets its food from the intestine. It can live without much oxygen. If you look at a stained Giardia under a microscope, it looks like a "smiley face"!
People often get Giardia by drinking unsafe water. This is the most common way it spreads. Eating contaminated food or soil can also cause infection. This is especially true in places with poor sanitation. Natural water sources can also get contaminated. This happens if animals graze near them.
Giardia infections happen all over the world. In the United States and Canada, Giardia is a common parasite. It is often found in children in day care centers. Hikers, family members, and people with weak immune systems can also get it. About 20,000 cases are reported each year in the United States.
Contents
Understanding the Giardia Life Cycle

Giardia has two main forms during its life. One form is called a trophozoite. This is a pear-shaped cell that moves around. It only lives inside the small intestine of a host. Trophozoites swim in the intestine's mucus. They then stick to the intestinal lining.
Once attached, trophozoites multiply by splitting in two. They can make more trophozoites or turn into a different form called a cyst. Cysts are passed out of the host's body in their poop. Giardia cysts are very tough. They can survive in the environment for weeks or even months. They need to stay moist to survive.
Cysts stay inactive until an animal or person swallows them. Inside the new host, the cyst changes. It releases two trophozoites. These new trophozoites then attach to the intestine. This starts the whole life cycle again.
Where Giardia Lives and Spreads
Giardia cysts can live for a long time in cold water. This means they can be found in contaminated wells and water systems. They are common in still water sources. These include ponds, storm drains, and even clean-looking mountain streams.
Cysts can also be on surfaces, soil, or food. This happens if they are touched by poop from infected humans or animals. City water reservoirs can also have cysts. Cysts are hard to remove with normal water treatment. They can resist chlorine and other methods.
Animals can also spread Giardia to humans. This is called zoonotic transmission. So, campers or swimmers in contaminated lakes might get sick. This is why giardiasis is sometimes called "beaver fever." Beavers can build dams that create still water.
Besides water, Giardia can spread from poop to mouth. This often happens in day-care centers. Children might not wash their hands well. People who work with children are also at risk. Family members of infected people can also get it. Not everyone who gets Giardia feels sick. Many people can carry the parasite without knowing it.
Giardia infects humans, but it also infects many animals. It is common in cats, dogs, and birds. It can infect dozens of other mammals. These include cattle, sheep, and goats.
Infected cats can be easily treated. Lambs usually just lose weight. But in calves, Giardia can be deadly. It often does not respond to medicines. Calves can also carry the parasite without showing symptoms. This parasite is very dangerous for chinchillas. So, they must always have safe water.
Dogs have a high infection rate. About 30% of puppies under one year old in kennels get infected. Puppies get it more often than adult dogs. Infected dogs can be kept separate and treated. Or, all dogs in a kennel can be treated together. Kennels should be cleaned with bleach. Grass areas used for exercise can stay contaminated for at least a month. This is because cysts can live a long time outside. To prevent spread, infected dogs should be kept separate for at least 20 days. Also, always provide clean water.
How Scientists Study Giardia
Scientists like Dr. Frances Gillin have studied Giardia in the lab. They learned how the parasite changes forms. They found out what signals in the host's body trigger these changes. They also discovered how Giardia avoids the body's defenses.
One way Giardia hides is by changing its surface proteins. This makes it hard for the immune system to find and fight the parasite. This is called antigenic variation. Dr. Gillin's work helps explain why Giardia infections can last a long time. It also shows why they can come back. Learning about Giardia's biology helps scientists. It helps them find better ways to prevent and treat infections.
In 2008, scientists discovered how Giardia uses a special process. It helps the parasite switch its surface proteins. This helps it avoid the host's immune system. This discovery was made by Dr. Hugo Lujan's team in Argentina.
The main meeting for Giardia research is the "International Giardia and Cryptosporidium Conference" (IGCC). Scientists share their new findings there.
A Brief History of Giardia Discovery


The first likely mention of Giardia was in 1681. Antonie van Leeuwenhoek described tiny "animalcules" in his own poop. They looked like Giardia.
The next known description was in 1859. A Czech doctor named Vilém Dušan Lambl wrote about the trophozoite form. He saw it in the poop of a child patient. Lambl named it Cercomonas intestinalis.
In 1888, Raphaël Blanchard renamed the parasite Lamblia intestinalis. He did this to honor Lambl. Then, in 1915, Charles Wardell Stiles gave it the name Giardia lamblia. This honored both Lambl and Professor Alfred Mathieu Giard. In 1921, Charles E. Simon drew detailed pictures of the parasite.
Images for kids
-
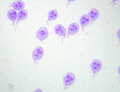
Giardia trophozoites stained with Giemsa; 100x magnification.
-
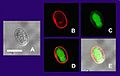
Multiple views of a Giardia lamblia cyst imaged by confocal microscopy. Bar = 10 micrometers
(A) Cyst imaged by transmission (differential interference contrast).
(B) Cyst wall selectively imaged through use of fluorescent-labelled antibody.
(C) Cyst imaged through use of carboxy fluorescein diacetate, a viability stain.
(D) Composite image of (B) and (C).
(E) Composite image of (A), (B), and (C).

See also
 In Spanish: Giardia lamblia para niños
In Spanish: Giardia lamblia para niños
