
Coues' rice rat facts for kids
Quick facts for kids Coues' rice ratTemporal range: Late Pleistocene to Recent
|
|
|---|---|
 |
|
| Conservation status | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Rodentia |
| Family: | Cricetidae |
| Genus: | Oryzomys |
| Species: |
O. couesi
|
| Binomial name | |
| Oryzomys couesi (Alston, 1877)
|
|
 |
|
| Distribution of Oryzomys couesi (in red) and other species of Oryzomys. | |
| Script error: The function "autoWithCaption" does not exist. | |
| Synonyms | |
and see below. |
|

Script error: No such module "Check for conflicting parameters".
The Coues' rice rat (scientific name: Oryzomys couesi) is a type of rodent that loves water. It lives in wet places like marshes and swamps, but can also be found in drier forests and shrublands. These rats are found from southern Texas, through Mexico and Central America, all the way to northwestern Colombia.
Coues' rice rats are medium to large-sized rats, weighing about 43 to 82 grams (1.5 to 2.9 ounces). They have rough fur that is buff (light brown) to reddish on top, and white or buff underneath. Their back feet are a bit special, helping them live in water. They have 56 chromosomes. These rats are active at night and build their nests out of plants, often hanging them about 1 meter (3.3 feet) above the ground in reeds. They are excellent swimmers and divers, but can also climb plants. They eat both plants and animals, including seeds and insects. Females usually have about four babies after being pregnant for 21 to 28 days.
Scientists first described this species in 1877. For a long time, many similar rats were thought to be the same species as the Coues' rice rat. However, more recent studies, especially using DNA in 2010, suggest that what we call Oryzomys couesi might actually be several different species. Generally, the Coues' rice rat is common and not considered endangered. In some places, there are so many that they are even seen as pests.
Contents
What is the Coues' Rice Rat?
The Coues' rice rat belongs to a group of animals called the Oryzomys couesi group. This group includes the Coues' rice rat and at least six other species that live in specific areas. Another well-known member of the Oryzomys genus is the marsh rice rat (O. palustris), which lives in the United States.
All these rice rats are part of a larger group called Oryzomyini, which has over a hundred different species. They are also part of the Cricetidae family, which includes hundreds of other small rodents like hamsters and voles.
How Scientists Study Them
Edward Alston first described the Coues' rice rat in 1877. He named it Hesperomys couesi, honoring American naturalist Elliott Coues. Later, in 1893, Oldfield Thomas moved the species to the genus Oryzomys. He also noticed that the first specimens used to describe the species were actually from different types of rats!
Over the years, many scientists studied these rats. In 1918, Edward Alphonso Goldman grouped many forms into one species, Oryzomys couesi, with ten different types (subspecies). In 1960, Raymond Hall thought the Coues' rice rat was the same species as the marsh rice rat from the U.S.
However, in 1979, new studies in Texas showed that the Coues' rice rat and the marsh rice rat were actually different species. Since then, some other types of rice rats that were once grouped with Oryzomys couesi, like Oryzomys antillarum from Jamaica and Oryzomys peninsulae from Baja California, have been recognized as their own separate species again.
Scientists continue to study the Coues' rice rat using DNA to understand how different populations are related. A 2010 study found that the Coues' rice rat might actually be four different species. For example, rats from the Pacific coast of Mexico and Central America might be one species, and those from the eastern side (Texas to Nicaragua) might be another. There also seem to be unique species in Panama and Costa Rica. More research is needed to confirm these findings and give them proper names.
What Does the Coues' Rice Rat Look Like?
The Coues' rice rat is a medium to large-sized rat. Its fur is rough and can be buff (light brown) to reddish on its back. The fur gets lighter on its sides and cheeks, and darker on its rump and face. Its belly is usually white or buff. Compared to the marsh rice rat, its fur is shorter, brighter, and more colorful.
It has a blunt snout and moderately large eyes that can show a reddish glow. Its small ears are black on the outside and covered with short, gray or reddish hairs on the inside. The tail is long, dark brown on top, and lighter brown or white underneath. Its feet are long and strong.
Special Features for Water Life
The Coues' rice rat has some special features that help it live in water:
- Its back feet have small webs between the toes, like a duck's foot.
- The pads on its hindfeet are smaller.
- The tufts of hair around its toes are reduced.
These features are common in animals that spend a lot of time in the water.
Inside the Body
Like many other rodents in its family, the Coues' rice rat does not have a gall bladder. It has 56 chromosomes.
Skull and Teeth
The skull of the Coues' rice rat has some unique features. For example, the cheekbones (zygomatic arches) are wide. The teeth are low-crowned and have many extra ridges, which is common for rice rats. They have 16 teeth in total: one incisor and three molars on each side of their upper and lower jaws.
How Does the Coues' Rice Rat Live?
The Coues' rice rat lives in many different places, from southern Texas down to Colombia. It prefers wet areas like marshes and small streams. However, it can also live in forests and shrublands as long as there's enough cover. You might even find them in sugarcane and rice fields. They can live from sea level up to about 2,300 meters (7,500 feet) high.
Behavior and Diet
These rats spend a lot of time in the water and are excellent swimmers and divers. They use their tails to help them move in the water. They can also climb plants very well. When they feel threatened, they often jump into the water and swim away.
Coues' rice rats are mostly active at night. They build round nests out of plants, often hanging them about 1 meter (3.3 feet) above the water or ground. Larger rats tend to build larger nests.
They eat a variety of foods, including both plants and animals. Their diet can include seeds, green plant parts, small fish, crabs, snails, and insects like ants and beetles.
Reproduction
Coues' rice rats can breed all year round. After being pregnant for 21 to 28 days, a female usually gives birth to about four young. The young rats can start having their own babies when they are about seven weeks old.
Health and Predators
Like many animals, Coues' rice rats can get parasites like ticks, mites, fleas, and worms. They can also carry two types of hantaviruses, but these viruses have not been linked to human illnesses. Snakes like the Boa constrictor are known to hunt Coues' rice rats, especially on islands like Cozumel.
Is the Coues' Rice Rat Endangered?
The IUCN says the Coues' rice rat is a species of "Least Concern". This means it's not currently endangered. It lives in many places, is quite common, and can adapt to different habitats. It also lives in many protected areas.
However, some local populations can be threatened. For example, if their wetland homes are drained, they might lose their habitat. In some areas, there are so many Coues' rice rats that people consider them a pest.
In Texas, where their habitat is very limited, they are listed as a threatened species because of habitat loss. A study in 2001 even predicted that climate change could cause the Texas population to disappear. The population on Cozumel island has also decreased a lot since the mid-1980s, possibly due to habitat changes and predators that were brought to the island.
Images for kids
-
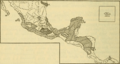
Distribution of Oryzomys couesi and related species according to Goldman (1918).
-
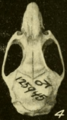
Skull of Oryzomys from Michoacán, Mexico (seen from above).
-
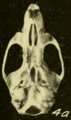
Skull of Oryzomys from Michoacán, Mexico (seen from below).
-
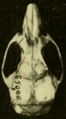
Skull of Oryzomys from Brownsville, Texas (seen from above).
-
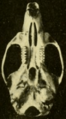
Skull of Oryzomys from Brownsville, Texas (seen from below).
-
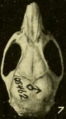
Skull of Oryzomys from Cozumel, Mexico (seen from above).
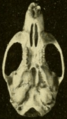
-
Skull of Oryzomys from Cozumel, Mexico (seen from below).
-
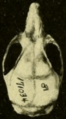
Skull of Oryzomys from Gatún, Panama (seen from above).
-
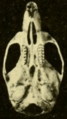
Skull of Oryzomys from Gatún, Panama (seen from below).
-
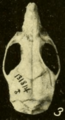
Skull of Oryzomys from Yaruca, Honduras (seen from above).
-
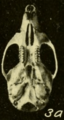
Skull of Oryzomys from Yaruca, Honduras (seen from below).











See also
 In Spanish: Rata arrocera de Coues para niños
In Spanish: Rata arrocera de Coues para niños
