
Fish scale facts for kids
A fish scale is a small, tough plate that grows from a fish's skin. Most fish are covered with these scales, which protect them. Scales can also help fish hide by reflecting light and using colors. They might even help fish swim better. The word "scale" comes from an old French word meaning "shell" or "husk."
Fish scales come in many different sizes, shapes, and structures. Some fish, like shrimpfish and boxfish, have strong, rigid armor-like scales. Others, like eels and anglerfish, have tiny scales or no scales at all. The shape of a scale can even help scientists figure out what kind of fish it came from.
Most bony fish have cycloid scales (like salmon and carp), ctenoid scales (like perch), or ganoid scales (like sturgeons and gars). Cartilaginous fish, such as sharks and rays, have placoid scales. Some fish have scutes, which are like bony plates, and others have no scales on some or all of their skin.
Fish scales are part of a fish's outer skin system. They grow from the inner skin layer, which is different from how reptile scales grow. The same genes that help teeth and hair grow in mammals are also involved in scale development. The placoid scales of sharks are also called dermal denticles, and they are similar in structure to vertebrate teeth. Most fish also have a protective layer of mucus, or slime, on their skin.
Contents
Thelodont Scales

The bony scales of thelodonts, which are very common fossil fish, are well understood. These fish would grow and shed their scales throughout their lives. After they died, the scales would quickly separate from their bodies.
Bone is strong and often preserves details well, so scientists can study the scales' tissues and how they grew. These scales have a "crown" made of dentine (a tooth-like material) with a shiny, enamel-like top surface. The base of the scale is made of bone and sometimes had structures to help it stay attached to the fish.
Scientists have found five types of bone growth in these scales, which might mean there were five main groups of thelodonts. Each type of scale seems to look like the scales of more modern fish groups, suggesting that thelodonts might be ancestors to later fish.
However, using only scale shape to identify species can be tricky. The shape of scales can change a lot depending on where they are on the fish's body. Also, the same scale shape might be found on two different species, which can be confusing.
Studying the shape and tissue of thelodont scales is the main way to understand their variety and tell different species apart. Even though similar features can evolve separately in different species, this method is still very useful. By comparing them to modern sharks, scientists have learned that thelodont scales worked similarly to modern cartilaginous fish scales. This has helped them understand the ancient fish's environment and lifestyle.
Cosmoid Scales

Cosmoid scales are only found on ancient lobe-finned fish, including some early lungfish and the living coelacanth (though its scales are slightly different). They likely developed from placoid scales joining together. The inside of these scales is made of dense, layered bone called isopedine. On top of this is a layer of spongy bone with blood vessels, then a complex dentine-like layer called cosmine, and finally a hard, shiny outer coating. The very top surface is keratin, a tough protein. Cosmoid scales grow larger as the layered bone inside them grows.
Elasmoid Scales

Elasmoid scales are thin, overlapping scales made of a layer of dense, layered bone called isopedine. Above this, there's usually a layer of small bumps, also made of bone. The dentine layer found in the first lobe-finned fish is usually smaller or completely missing in modern fish like lungfish and coelacanths.
Elasmoid scales have appeared many times during fish evolution. They are found in some lobe-finned fish, including all living and some extinct lungfish, as well as coelacanths, which have modified cosmoid scales that are thinner and lack cosmine. They are also present in some fish that are ancestors to four-legged animals, like Eusthenopteron, and in modern bony fish (teleosts), whose cycloid and ctenoid scales are the least hardened elasmoid scales.
Ganoid Scales
_(3149758934).jpg)
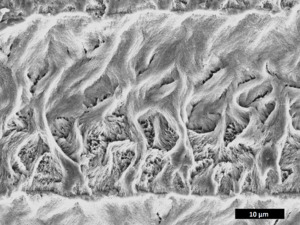
Ganoid scales are found on fish like sturgeons, paddlefish, gars, bowfin, and bichirs. They developed from cosmoid scales and often have jagged edges. They are covered with a hard, enamel-like layer instead of cosmine, and a layer of a mineral called ganoine instead of vitrodentine.
Ganoine is a special part of ganoid scales. It's a glassy, often layered mineral tissue that covers the scales, and sometimes the skull bones and fin rays of some ray-finned fish like gars, bichirs, and coelacanths. It's made of tiny rod-like crystals. Ganoine is an ancient feature of ray-finned fish, found even on the scales of early ray-finned fish like Cheirolepis. Some scientists think ganoine is similar to tooth enamel in vertebrates.
 |
Ganoid scales of the extinct Carboniferous fish, Amblypterus striatus. (a) shows the outer surface of four of the scales, and (b) shows the inner surface of two of the scales. Each of the diamond-shaped ganoid scales of Amblypterus has a ridge on the inner surface which has a projecting peg. This peg fits into a notch in the next scale, similar to how tiles are pegged together on a roof. |  |
Most ganoid scales are diamond-shaped and connect with peg-and-socket joints. They are usually thick and fit together like a jigsaw puzzle rather than overlapping. This makes ganoid scales almost impossible to break through, offering excellent protection against predators.
-

The alligator gar has tough armor made of diamond-shaped ganoid scales
-

The sturgeon has rows of ganoid scales that are enlarged into scute-like armor plates
-

The ganoid scales on a bowfin are smaller and look like cycloid scales

.png)


In sturgeons, the scales are much larger and form armor plates along their sides and back. In the bowfin, the scales are much thinner and look like cycloid scales.
-

Earrings made from the ganoid scales of an alligator gar
-

Fossil of a primitive rayfin with ganoid scales
-

Ganoid scales on a fossilized Lepidotes, from about 130 million years ago


.jpg)
Native Americans and people in the Caribbean used the tough ganoid scales of the alligator gar for arrowheads, chest plates, and to cover plows. Today, these scales are sometimes used to make jewelry.
Leptoid Scales
Leptoid scales are found on more advanced bony fish called teleosts. The outer part of these scales spreads out with bony ridges, while the inner part has criss-crossing fibrous tissue. Leptoid scales are thinner and more see-through than other types of scales. They don't have the hard enamel-like or dentine layers. As the fish grows, new scales are added in layers, like tree rings.
Leptoid scales overlap like roof tiles, from head to tail. This makes them more flexible than cosmoid and ganoid scales. This arrangement helps water flow smoothly over the fish's body, reducing drag. The scales of some fish show bands of uneven growth called annuli (singular annulus). These bands can be used to figure out the fish's age.
Leptoid scales come in two main forms: cycloid and ctenoid.
Cycloid Scales
Cycloid scales are smooth and round, with a smooth outer edge. They are most common on fish with soft fin rays, such as salmon and carp.
 |
.jpg) |
|
|
|


|
|
|







Ctenoid Scales
Ctenoid scales are similar to cycloid scales, but they have small teeth or spines called ctenii along their outer edges. Because of these teeth, the scales feel rough. They are usually found on fish with spiny fin rays, such as perch-like fish. These scales have almost no bone; they are made of a surface layer with minerals and a deeper layer mostly of collagen (a type of protein). The enamel layer found on other scale types is reduced to small ridges and ctenii.


 |
 |
|
|
|

_by_M._L._Nievera_(colored).png)
|
|
|





Ctenoid scales, like other skin structures, start developing from special skin areas. Their unique cell development makes them different from other skin parts. Development begins near the tail fin, along the fish's lateral line. First, cells called fibroblasts gather between the outer and inner skin layers. Then, collagen fibers (a type of protein) start to organize in the inner skin layer, which leads to the scales hardening with minerals. The scales first grow in circumference, then in thickness as overlapping layers harden together.
Ctenoid scales can be divided into three types:
- Crenate scales: These have wavy edges with bumps and dips.
- Spinoid scales: These have spines that are part of the scale itself.
- True ctenoid scales: These have spines that are separate structures on the scale.
Most ray-finned fish have ctenoid scales. Some flatfish have ctenoid scales on their eyed side and cycloid scales on their blind side. Other species have ctenoid scales in males and cycloid scales in females.
Reflection



Many teleost fish have very reflective scales that look like silvery glass. This silvering is common in fish that live in the open ocean, especially in the top 100 meters of water. Silvering helps fish become almost invisible by making their bodies highly reflective. In the middle depths of the ocean, light comes from above, so a mirror-like body oriented vertically makes fish invisible from the side.
The marine hatchetfish is very flat from side to side, only a few millimeters thick, and its body is so silvery it looks like aluminum foil. These "mirrors" are made of tiny structures, like stacks of 5 to 10 guanine crystals. These crystals are spaced just right to reflect almost 100% of the light. In the deep waters where hatchetfish live, only blue light reaches them. So, mirrors spaced 125 nanometers apart (a very tiny distance) provide good camouflage.
Most fish in the upper ocean use silvering to hide. Fish like herring, which live in shallower water, need to reflect a mix of light colors. So, their crystal stacks have different spacings. For fish with rounded bodies, flat mirrors wouldn't work well. Instead, they have many small reflectors, all angled vertically, to create the overall mirror effect.
Fish scales with these reflective properties are sometimes used in cosmetics to give makeup and lipstick a shimmering effect.
Placoid Scales

Placoid scales (pointed, tooth-shaped) are found on cartilaginous fish: sharks, rays, and chimaeras. They are also called dermal denticles (which means "skin teeth"). Placoid scales are similar in structure to vertebrate teeth. They have a soft center (pulp cavity) with blood vessels, surrounded by a cone-shaped layer of dentine. All of this sits on a rectangular base plate in the inner skin layer. The outermost layer is a hard, enamel-like substance. Placoid scales don't grow larger; instead, more scales are added as the fish grows.
Similar scales can also be found under the head of the denticle herring. Rays and chimaeras have much less scale coverage.
Shark Skin

Shark skin is almost completely covered by small placoid scales. These scales have small spines that feel rough when you rub them backward. But when the shark moves forward, the spines flatten, creating tiny swirling water currents that reduce water resistance and make swimming more efficient and quieter than for bony fish. This also helps prevent things from sticking to the shark's skin, like barnacles.
All dermal denticles have a soft center with nerves and blood vessels connected to the inner skin layer, which supplies them with mucus. Denticles have small ridges that stick out from the surface. Under a microscope, these ridges can look like hooks or lines. The shape of these ridges depends on the type of shark. Generally, there are two main shapes: ridges that run sideways along the shark, parallel to the water flow, or smooth scales with hook-like ridges that curl backward. Both shapes help create a swirling layer of water near the shark's skin, pushing the smooth water flow further away from the shark.
Unlike bony fish, sharks have a complex inner "corset" made of flexible protein fibers arranged in a spiral pattern around their body. This corset acts like an outer skeleton, giving their swimming muscles something to attach to and saving energy. Depending on where they are on the body, these placoid scales can be flexible and can stand up, changing the angle at which water hits them. These scales also have small ridges that are lined up with the water flow. These ridges reduce the drag force on the shark's skin by pushing the swirling water further away from the surface, preventing high-speed cross-stream flow.
Scale Shape and Drag Reduction
The general shape of the scales varies, but all have three parts: the crown, the neck, and the base. How flexible a scale is depends on the size of its base. Scales with smaller bases are more flexible and less firmly attached to the skin. On the crown of fast-swimming sharks, there are parallel ridges that run from front to back.
Scientists have found that the base of the denticle doesn't touch the water flow. However, the crown and neck of the denticles are key to creating the swirling water currents near the skin's surface. Since denticles come in many shapes and sizes, not all shapes create the same type of swirling flow. In one experiment, artificial shark denticles with a crescent shape were tested in a water tank. The results showed that the surface with denticles reduced drag by 10% compared to a smooth surface. This drag reduction happened because the swirling water currents got trapped between the denticles, creating a "cushion" against the smooth water flow. Other research confirmed these findings, showing that more practical, low-profile shapes (like trapezoidal or semi-circular troughs) were less effective but still reduced drag by 6-7%.
Reducing Drag

Sharks reduce drag and the energy they use to move in several ways. Drag comes from two main sources:
- Pressure drag: This is caused by the difference in pressure between the front and back of the shark as it pushes water aside to move forward. It increases with smoother water flow.
- Frictional drag: This results from the water rubbing against the shark's skin. It changes depending on how the water layer next to the skin behaves.
The small ridges on shark scales prevent swirling water currents from forming too close to the surface. The mechanism is complex, but basically, the ridges are too small for the swirling water to fit into their valleys. This pushes the swirling water further away from the surface, so it only interacts with the tips of the ridges, which is a very small area. This greatly reduces the transfer of movement that causes drag. It also reduces sideways water movements, which further helps reduce drag.
Recent studies show that the water layer near the shark's skin goes through a cycle of thickening and then breaking down into swirling currents before collapsing. This system is self-regulating: the swirling currents build up and then quickly break apart into organized patterns of "hairpin" vortices that lift off the wall. These lifting vortices push the water layer away from the shark's surface, which reduces the overall drag the fish experiences.
The rough, sandpaper-like texture of shark and ray skin, combined with its toughness, has made it valuable as a source of leather called shagreen. Historically, shark shagreen was used for sword grips. The rough texture is also used in Japanese cooking to make graters called oroshiki, by attaching pieces of shark skin to wooden boards. The small scales grate food very finely.
Technical Uses
.jpg)
In the marine industry, there's a huge need for surfaces that prevent things from sticking to them. This is called anti-fouling, and it stops barnacles, algae, and other growths from covering boat hulls. Dermal denticles are a very promising area of research for this because sharks are one of the only fish that don't get covered in these growths. Studies by the U.S. Navy have shown that if a material that imitates shark skin could be made, it could save military vessels up to 45% in fuel.
There are many examples of materials and surfaces that copy the structure of aquatic animals, including sharks. These are designed to help things move more efficiently through liquids like air, water, and oil.
Surfaces that mimic shark skin have also been used to prevent microorganisms and algae from coating the hulls of submarines and ships. One type is sold as "sharklet."
Many new ways to copy shark skin involve using a type of silicone called polydimethylsiloxane (PDMS) to create a mold. Usually, a flat piece of shark skin is covered with PDMS to make a mold, and then more PDMS is poured into that mold to create a shark skin replica. This method has been used to create a surface that is extremely water-repellent, showing the lotus effect (where water beads up and rolls off, taking dirt with it). One study found that these artificial surfaces reduced drag by up to 9%, and with a flapping motion, drag reduction reached 12.3%.
Besides marine uses, the aerospace industry can also benefit from these designs. Scientists have created computer models of shark denticles with different shapes, like low and high-profile vortex generators. These models were designed and analyzed to see how applying denticle-like structures to airplane wings would affect them. During simulations, it was noted that the samples changed how the wings reacted at low and high angles of attack (the angle at which air hits the wing). The low-profile vortex generators performed 323% better than current smooth wing structures. This improvement is due to a "separation bubble" in the denticles' wake and swirling air currents that restore lost momentum in the air layer next to the wing.
Scutes
_Waka,_Japan.jpeg)
Scutes are similar to scales and have the same job of protecting the fish. However, unlike fish scales, which form from the outer skin layer, scutes form in the lower, blood-rich layer of the skin. The outer part of the scute is a tough, horny layer, which looks like a scale.
The word "scute" comes from Latin for "shield." Scutes can be:
- An external, shield-like bony plate.
- A modified, thickened scale that often has a ridge or is spiny.
- A projecting, modified (rough and strongly ridged) scale, usually found along the lateral line (a sensory line on the fish's side) or on the narrow part before the tail, forming tail keels, or along the belly.
Some fish, like pineconefish, are completely or partly covered in scutes. River herrings and threadfins have a row of scutes on their belly, which are scales with raised, sharp points used for protection. Some jacks have a row of scutes along their lateral line.
Scale Development
Scales usually appear late in a fish's development. For example, in zebrafish, it takes 30 days after fertilization for the different layers needed to start forming scales to develop and organize. First, a type of embryonic tissue needs to become solid, then the shape of the scales starts to form, and finally, the cells specialize, completing the process.
Modified Scales




Different groups of fish have developed modified scales to serve various purposes.
- Almost all fish have a lateral line, which is a system of sensors that detect water movements. In bony fish, the scales along the lateral line have central pores that allow water to reach these sensory cells.
- The dorsal fin spines of dogfish sharks and chimaeras, the stinging tail spines of stingrays, and the "saw" teeth of sawfish and sawsharks are all fused and modified placoid scales.
- Surgeonfish have a sharp, scalpel-like blade, which is a modified scale, on each side just before their tail.
- Some herrings, anchovies, and halfbeaks have deciduous scales, which are easily shed. This helps them escape from predators.
- Male Percina darters have a row of enlarged caducous scales between their pelvic fins and their anus.
- Porcupine fish have scales modified into large external spines.
- Pufferfish have thinner, more hidden spines than porcupine fish. These spines only become visible when the fish puffs up. Unlike porcupine fish, these spines are not modified scales, but they develop using the same genes that produce feathers and hairs in other vertebrates.
-
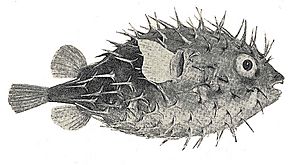
Porcupine fish have scales modified into spines
-

Pufferfish spines are not modified scales, but developed by an independent gene network


Fish Without Scales
-

Mandarinfish lack scales and protect themselves with a layer of smelly and bitter slime.

Fish without scales usually have other ways to protect themselves, such as tough, leathery skin or bony plates.
- Jawless fish (lampreys and hagfish) have smooth skin without scales or dermal bone. Lampreys have tough, leathery skin for protection. Hagfish produce a lot of slime if they feel threatened. They can tie themselves in a knot to scrape off the slime and escape from a predator.
- Most eels don't have scales, though some species have tiny, smooth cycloid scales.
- Most catfish don't have scales, but several families have body armor in the form of bony plates or scutes.
- Mandarinfish don't have scales and have a layer of smelly, bitter slime that prevents disease and likely discourages predators. Their bright colors might be a warning to others.
- Anglerfish have loose, thin skin often covered with fine, forked skin prickles or bumps, but they don't have regular scales. They rely on camouflage to avoid predators, and their loose skin makes it hard for predators to grab them.
Many groups of bony fish, including pipefish, seahorses, boxfish, poachers, and several families of sticklebacks, have developed external bony plates as protective armor against predators.
- Seahorses don't have scales. Instead, they have thin skin stretched over bony plate armor arranged in rings along their bodies.
- In boxfish, the plates fuse together to form a rigid shell that covers their entire body. These bony plates are not modified scales; they are skin that has turned into bone. Because of this heavy armor, boxfish move slowly, but few other fish can eat adult boxfish.
 |
|
|
|
|
-

Boxfish have plates of hardened skin fused together to form a rigid shell
-

Seahorses have thin skin stretched over bony plates arranged in rings


Some fish, like hoki and swordfish, are born with scales but lose them as they grow older.
Filefish have rough, non-overlapping scales with small spikes, which is why they are called filefish. Some filefish look scaleless because their scales are so tiny.
Tuna only have noticeable scales along their lateral line and in a protective band of thickened, enlarged scales around their shoulders called a corselet. Over most of their body, tuna have scales so small that they seem scaleless to a quick look.
-

Some filefish appear scaleless because their scales are so small.
-

To a quick look, tuna seem mostly without scales, but they do have them.


Religious Rules
In the Bible, the Book of Leviticus says that "of all that are in the waters... in the seas, and in the rivers," those that do not have both fins and scales "shall be an abomination unto you" and cannot be eaten. This means all water invertebrates are forbidden, as well as any fish that lack scales (there don't seem to be fish that lack fins).
According to Jewish law (Torah and Talmud), for a fish to be considered kosher (allowed to eat), it must have scales and fins. The definition of "scale" here is different from what biologists use. For a fish to be kosher, its scales must be visible to the eye, present in the adult fish, and easily removed from the skin by hand or with a scaling knife. If the scales can't be removed without tearing the skin, the fish is not kosher. So, carp and salmon are kosher. However, a shark, whose scales are microscopic, a sturgeon, whose scutes can't be easily removed, and swordfish, which lose their scales as adults, are all not kosher. Other non-kosher fish include catfish, eels, Pacific cod, snake mackerels, and puffer fish.
Lepidophagy

Lepidophagy (an ancient Greek word meaning "scale-eating") is a special feeding behavior in fish where they eat the scales of other fish. This behavior has developed independently in at least five freshwater fish families and seven marine fish families.
Fish scales can be nutritious. They contain a skin part and a layer of protein-rich mucus, in addition to layers of keratin and enamel. They are also a good source of calcium phosphate. However, the energy a fish uses to attack another fish versus the amount of scales it gets limits the size of scale-eating fish. They are usually much smaller than their prey. Scale-eating often develops when food is scarce or in extreme environments. Eating scales and the skin around them provides protein-rich nutrients that might not be available elsewhere.
Fish jaws usually have bilateral symmetry, meaning they are the same on both sides. An exception is the scale-eating cichlid Perissodus microlepis. This fish has two distinct jaw shapes. One shape has its jaw twisted to the left, making it easier to eat scales from the right side of its victim. The other shape has its jaw twisted to the right, making it easier to eat scales from the left side of its victim. The number of each jaw shape in a population is controlled by how common each shape is.
See also
 In Spanish: Escama de pez para niños
In Spanish: Escama de pez para niños
