
Green Mountain maidenhair fern facts for kids
Quick facts for kids Green Mountain maidenhair |
|
|---|---|
 |
|
| Adiantum viridimontanum growing in a dunite roadcut | |
| Conservation status | |
| Scientific classification |
|
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Class: | Polypodiopsida |
| Order: | Polypodiales |
| Family: | Pteridaceae |
| Genus: | Adiantum |
| Species: |
A. viridimontanum
|
| Binomial name | |
| Adiantum viridimontanum C.A.Paris
|
|
| Script error: The function "autoWithCaption" does not exist. | |
| Synonyms | |
|
Adiantum pedatum subsp. viridimontanum (C.A.Paris) Fraser-Jenk. |
|

Script error: No such module "Check for conflicting parameters".
The Adiantum viridimontanum, also known as the Green Mountain maidenhair fern, is a very rare fern. You can only find it growing on special rocks called serpentine in New England and Eastern Canada.
This fern has unique leaves that look like fingers. These "fingers" are actually parts of one large compound leaf. They grow from a dark, shiny central stalk. The fern's "fingers" can hang down or stand up straight. This depends on whether the fern grows in the shade or in sunlight. Like other ferns in its group, it carries its Spores under rolled-up flaps on the edges of its leaves.
For a long time, scientists thought this fern was the same as the western maidenhair fern. But in 1991, a botanist named Cathy Paris discovered it was a new species. She named it A. viridimontanum after the Green Mountains in Vermont, where she found it. Since then, it has also been found in Quebec and Maine.
Contents
About the Green Mountain Maidenhair Fern
What Does It Look Like?
The Green Mountain maidenhair fern is a medium-sized plant. It can grow up to about 2 feet (60 cm) wide and 1 to 2 feet (30 to 60 cm) tall. Its leaves, called fronds, can be from 30 to 75 cm (12 to 30 in) long.
The fronds of this fern look like they fork into two main branches. These branches curve outwards. Many smaller leaf sections, called pinnae, grow from the outer side of these curves. These pinnae are further divided into even smaller parts. The whole leaf looks like a hand with finger-like segments.
The fern has a rhizome (an underground stem) that is about 2 to 3.5 mm wide. The rhizome and the leaf stalk (called the stipe) have bronze-colored scales. The stipe and the main leaf stalk (rachis) are dark brown or purple.
The smallest parts of the leaf are called "ultimate segments." These segments are long and triangular. They are bright green to bluish-green. When the fern is young, its leaves are smooth and shed water easily. On the underside of the leaf, these segments have special flaps called false indusia.
How Does It Make Spores?
The fern's sporangia (where spores are made) are found under these false indusia. This is a special feature of all Adiantum ferns. The spores are round and yellow. They are about 41 to 58 micrometers (μm) wide. Spores are usually ready in the summer and fall.
How Is It Different from Other Ferns?
It can be tricky to tell the Green Mountain maidenhair fern apart from its close relatives. These are A. pedatum and A. aleuticum. No single feature makes it completely different from the others.
However, you can often tell it apart from A. pedatum by the shape of its smallest leaf parts. Also, A. viridimontanum can grow in both sun and shade, while A. pedatum prefers only shade.
To tell it apart from A. aleuticum, look at the stalks of the smallest leaf parts. They are longer in A. viridimontanum. Its false indusia are also longer. The spores of A. viridimontanum are also generally larger.
Where the Green Mountain Maidenhair Fern Lives

The Green Mountain maidenhair fern is found in only a few places. These include New England in the United States and Quebec in Canada. Many of its locations are in the Missisquoi Valley in the northern Green Mountains of Vermont. This is how it got its common name. It is also found in southern Quebec and at one spot in coastal Maine.
This fern loves sunny, disturbed areas with thin soil. It grows on dunite and other ultramafic rocks. You can often find it on talus slopes, along road cuts, and even in old asbestos mines. Human activities like mining and road building have helped create these open, sunny spots. In nature, falling rocks and erosion also help keep its habitat open.
This fern is one of only a few plant species that grow only on serpentine rock in eastern North America.
Life Cycle and Conservation
How Does It Reproduce?
The Green Mountain maidenhair fern mostly reproduces by growing new branches from its underground stem. It does not rely much on sexual reproduction through spores. While spores can be carried by the wind, most fall close to the parent plant.
For a spore to grow into a new fern, it needs to land in just the right spot. This means bright sunlight and thin serpentine soil. These conditions are often found in places that have been recently disturbed. This allows the fern to grow in new areas where other plants might not survive. The ferns seem to live a long time, which helps keep their populations stable even if new plants don't grow very often.
Why Is It Important to Protect This Fern?
Scientists don't know much about the fern's role in the ecosystem. Ferns generally have natural defenses against animals that might try to eat them. This fern is not known to be threatened by any specific animal or disease.
The Green Mountain maidenhair fern is considered "globally vulnerable." This means it is at risk of disappearing. Its main problem is that it only grows in very specific places, like serpentine cliffs and rocky slopes.
These areas are not usually valuable for human use. The biggest threats to the fern are the expansion of asbestos mining or other activities in old mine sites. Road construction could also harm some populations. However, the fern can sometimes grow well in disturbed serpentine areas. So far, no non-native plants are known to be invading its habitats. It is also harder to grow this fern in gardens compared to its relatives.
Images for kids
-

Adiantum viridimontanum growing in a dunite roadcut
-
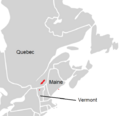
Natural distribution of A. viridimontanum within eastern Canada and the northeastern United States
