
Fission yeast facts for kids
Quick facts for kids Schizosaccharomyces pombe |
|
|---|---|
 |
|
| Scientific classification | |
| Kingdom: | |
| Division: | |
| Class: |
Schizosaccharomycetes
|
| Order: |
Schizosaccharomycetales
|
| Family: |
Schizosaccharomycetaceae
|
| Genus: |
Schizosaccharomyces
|
| Species: |
S. pombe
|
| Binomial name | |
| Schizosaccharomyces pombe Lindner (1893)
|
|
Schizosaccharomyces pombe, also known as "fission yeast", is a type of yeast. People have used it for a long time in making traditional beer. It is also very important in science as a "model organism". This means scientists study it to learn about how molecules and cells work.
Fission yeast is a single-celled eukaryote. Its cells are shaped like tiny rods. They are usually about 3 to 4 micrometres wide and 7 to 14 micrometres long. Its genome (all its genetic information) is about 14.1 million base pairs long. Scientists believe it has about 4,970 genes that make proteins. It also has at least 450 other types of RNA that do not make proteins.
These cells keep their rod shape by growing only at their ends. They divide into two equal "daughter cells" by splitting in the middle. This splitting process is called binary fission. Because of this clear division, fission yeast is a great tool for studying the cell cycle.
A scientist named Paul Lindner first found fission yeast in 1893. He found it in East African millet beer. The name pombe means "beer" in the Swahili language. Scientists started using it as a model in the 1950s. Urs Leupold used it to study genetics, and Murdoch Mitchison used it to study the cell cycle.
Paul Nurse, a famous fission yeast researcher, brought these two study areas together. He, along with Lee Hartwell and Tim Hunt, won the 2001 Nobel Prize in Physiology or Medicine. They won for their important work on how cells control their division.
The full genome of S. pombe was published in 2002. A group led by the Sanger Institute completed this work. It was the sixth eukaryotic model organism to have its full genome sequenced. Scientists who study S. pombe get help from the PomBase database. This has made the yeast even more useful for research. Scientists have found that about 70% of its genes are similar to human genes. Many of these are linked to human diseases. In 2006, scientists also mapped where almost all S. pombe proteins are located inside the cell. They used green fluorescent protein to do this.
Schizosaccharomyces pombe has also become important for studying how cells react to DNA damage. It helps us understand how cells fix their DNA. It also helps us learn about the process of DNA replication (when DNA makes copies of itself).
Scientists have found about 160 different types of S. pombe in nature. They have collected these from places like Europe, North and South America, and Asia. Most of these types were found on fruits like apples and grapes. They were also found in different alcoholic beverages, such as Brazilian Cachaça. S. pombe is also found in fermented tea, like kombucha. It is not clear if S. pombe is the main yeast that ferments these drinks or just a small part. We don't know much about where Schizosaccharomyces yeasts live naturally.
Contents
History of Fission Yeast Discovery
Schizosaccharomyces pombe was first found in 1893. A group in a German brewery laboratory was studying sediment in millet beer from East Africa. This sediment made the beer taste sour. The word "schizo" means "split" or "fission." It was used because this yeast divides by splitting. The word "pombe" was added because it means "beer" in Swahili.
The common S. pombe strains used today were found by Urs Leupold in 1946 and 1947. He got a culture from a yeast collection in Delft, The Netherlands. This culture had been put there by A. Osterwalder. Osterwalder had found it in 1924 in French wine. The culture Leupold used had different types of cells. Since then, scientists have made two big efforts to find S. pombe from fruits, nectar, or fermented drinks. One was in Sicily, and the other was in Brazil.
Where Fission Yeast Lives
The fission yeast S. pombe belongs to a large group of fungi called Ascomycota. These fungi are often found in tree sap, on plant roots, and in the soil around them. They also live on ripe and rotting fruits. Insects can carry them between different places. Many of these fungi live in a helpful way with plants. Others break down dead material. Some are even plant diseases.
The Schizosaccharomyces yeast is special among other yeasts. It has a unique cell wall that contains α-(1,3)-glucan. It also has very little chitin, which is common in other fungi. S. pombe can perform aerobic fermentation when there is a lot of sugar. It can also break down L-malic acid, which is a main acid in wine. This makes it different from other Saccharomyces yeasts.
Fission Yeast vs. Budding Yeast
Scientists study Schizosaccharomyces pombe and Saccharomyces cerevisiae (budding yeast) a lot. These two types of yeast separated from each common ancestor a very long time ago. They are both important tools in biology. Here are some ways they are different:
- S. cerevisiae has about 5,600 open reading frames (parts of genes that make proteins). S. pombe has about 5,070.
- Even though they have similar gene numbers, S. cerevisiae has only about 250 introns (non-coding parts of genes). S. pombe has nearly 5,000.
- S. cerevisiae has 16 chromosomes. S. pombe has only 3.
- S. cerevisiae is often diploid (has two sets of chromosomes). S. pombe is usually haploid (has one set of chromosomes).
- S. pombe stays in the G2 phase of the cell cycle for a long time. This means it controls when it enters the M phase (mitosis) very carefully. S. cerevisiae stays in the G1 phase for a long time.
- Both yeasts have genes similar to those in more complex organisms, but they don't always share these genes with each other. For example, S. pombe has genes for RNAi machinery like humans. S. cerevisiae does not.
- S. cerevisiae has small centromeres (parts of chromosomes important for cell division) that are 125 base pairs long. S. pombe has large, repeating centromeres that are 40 to 100 kilobases long. These are more like human centromeres.
Fission Yeast Life Cycle

Fission yeast is a single-celled fungus. It has a simple genome that scientists know well. It also grows very quickly. People have used it for a long time in brewing and baking. It is also used a lot in genetics research.
S. pombe cells are rod-shaped, about 3 micrometers wide. They grow longer only at their ends. After mitosis (when the nucleus divides), the cell divides. A wall, called a septum or cell plate, forms in the middle. This splits the cell into two equal parts.
The main steps of cell reproduction are:
- S (Synthetic) phase: Chromosomes make copies of themselves.
- M (Mitotic) phase: Chromosomes separate, the nucleus divides, and the cell splits.
- G1 phase: This is a gap between M and S phases.
- G2 phase: This is a gap between S and M phases.
In fission yeast, the G2 phase is quite long. The cell does not split until a new S phase begins. Fission yeast controls mitosis in ways similar to how animal cells do. It usually lives in a haploid state. If it runs out of food, cells of different "mating types" (P and M) can join together. They form a diploid zygote. This zygote then immediately goes through meiosis to make four haploid spores. When conditions get better, these spores grow into new haploid cells.
-

General features of the cell cycle.
-

The particular cell cycle of a fission yeast.
-
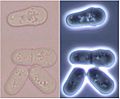
Division stages of Schizosaccharomyces under a microscope.


How Fission Yeast Cells Divide

Here's how fission yeast cells divide. The cell decides where it will split before a stage called anaphase. The spindle (shown in green in the picture) then moves. This makes sure the separated chromosomes are on opposite sides of where the cell will split.
How Cell Size is Controlled

In fission yeast, how much the cell grows affects when it moves from G2 to M phase. If a cell has a change in a gene called wee1, it enters mitosis when it is too small. This makes its G2 phase shorter. Also, cells with less food grow slowly. So, it takes them longer to double in size and divide. Low food levels also make the cell enter the cell cycle at a smaller size.
When S. pombe cells are under stress, like heat or a chemical called hydrogen peroxide, they start to "age." This means their division time gets longer, and they are more likely to die. Even with the wee1 gene change, cells are smaller but still take the same amount of time to go through the cell cycle. This is because smaller cells grow slower.
Scientists think a "spatial gradient" helps control cell size and when mitosis starts. A protein called Pom1 (green) is found on the cell's outer layer. It is most concentrated at the cell tips. Other proteins like Cdr2, Cdr1, and Wee1 are in nodes in the middle of the cell (blue and red dots).
- In small cells, the Pom1 protein reaches most of these nodes (blue dots). Pom1 stops Cdr2, which then stops Cdr1 from stopping Wee1. This allows Wee1 to stop cell division, so the cell does not enter mitosis.
- In long cells, the Pom1 protein does not reach the nodes (red dots). So, Cdr2 and Cdr1 stay active. They stop Wee1, which then allows cell division to start.
Mating-Type Switching
Fission yeast can switch its mating type. This happens during the S phase of the cell cycle. It uses a special process linked to how DNA copies itself. This was the first time scientists saw that the direction of DNA copying was needed for a cell to change its type. Studying this process led to finding special sites on the chromosome. These sites can stop or pause DNA copying. It also led to finding a new type of mark on one of the DNA copies. This mark helps control mating type. Work on this system has also helped us understand how heterochromatin (tightly packed DNA) forms and stays stable.
How Fission Yeast Responds to DNA Damage
Schizosaccharomyces pombe is a microorganism that can reproduce sexually. It does this when it doesn't have enough food. If S. pombe is exposed to hydrogen peroxide, which damages DNA, it strongly starts mating. It also forms meiotic spores. This suggests that meiosis (a type of cell division) and especially meiotic recombination (mixing of genetic material) might help repair DNA damage.
Scientists found that certain types of DNA damage can cause this recombination. This process needs an enzyme called uracil-DNA glycosylase. This enzyme removes damaged parts from the DNA. Based on this, scientists think that fixing certain DNA damage can start recombination in S. pombe. Other studies showed that problems with how DNA copies itself can also cause DNA damage. This damage then encourages meiotic recombination.
Fission Yeast as a Model System
Fission yeast has become a very important "model system" for scientists. It helps them study basic rules of cells. These rules can then help us understand more complex organisms, like humans. This single-celled eukaryote is not harmful. It is also easy to grow and change in the lab.
Fission yeast has one of the smallest numbers of genes among known eukaryotes. It has only three chromosomes. Many of the genes that control cell division and organization in fission yeast are also found in humans. Cell cycle control and division are vital for any cell to grow and develop. Scientists have studied fission yeast's similar genes a lot. This has led to many new discoveries in medicine.
Fission yeast is also good for watching cell division. This is because its cells are cylinder-shaped and divide by splitting in the middle. You can easily see this under a microscope. Fission yeast also grows very quickly, taking only 2 to 4 hours to reproduce. This makes it easy to study in the lab.
Because its genome is simple but similar to humans, it's easy to work with, and can be used for testing medicines, fission yeast helps a lot in medical and cell biology research. It is a great model for genetic studies.
Fission Yeast Genome
Schizosaccharomyces pombe is often used to study cell division and growth. This is because it has similar genetic parts to humans. These include proteins for heterochromatin, large origins of DNA copying, large centromeres, and how cells check for errors. It also helps us understand telomeres (ends of chromosomes), gene splicing, and other cell processes.
The full genome of S. pombe was sequenced in 2002. It was the sixth eukaryote genome to be sequenced. Scientists found about 4,979 genes on its three chromosomes. These chromosomes contain about 14 million base pairs of DNA. This DNA is in the cell's nucleus. After the first sequencing, other parts of the genes that were not sequenced before have now been mapped. You can find details about these genes on large databases like PomBase.
About 43% of the genes in the genome were found to have introns. Fission yeast does not have as many duplicated genes as budding yeast, only about 5%. This makes fission yeast a great model to study genes. Having many introns also means that one gene can make different types of proteins. This is similar to how human genes work.
About 81% of the three centromeres in fission yeast have been sequenced. Their lengths are 34, 65, and 110 kilobases. This is much longer than the centromeres of budding yeast. There is also a very high similarity (97%) in a specific region of the centromere. These long and similar centromeres make fission yeast a good model to study cell division in humans.
PomBase reports that over 69% of S. pombe protein-coding genes have similar genes in humans. More than 500 of these are linked to human diseases. This makes S. pombe a great system to study human genes and disease pathways. It is especially useful for understanding cell cycle and DNA checkpoint systems.
Genetic Diversity
Scientists have studied the variety and evolution of fission yeast. They looked at 161 types of Schizosaccharomyces pombe from 20 countries. Their research showed that all these types came from a common ancestor that lived about 2,300 years ago. The study also found 57 types of fission yeast that were very different from each other. All 57 types could grow on the same basic food as the main reference type.
Some studies suggest that the genetic variety of fission yeast is a bit less than that of budding yeast. There are only small differences in how S. pombe grows in different places. Also, the amount of visible differences in S. pombe is less than what is seen in S. cerevisiae. Since most types of fission yeast were found in brewed drinks, we don't know much about how they spread naturally.
Fission Yeast as a Biomedical Tool
There are some limits to using fission yeast as a model system. One is its ability to resist many medicines. This resistance happens because the yeast makes special "drug efflux pumps." These pumps push medicines out of the cell.
Paul Nurse and his team have created new S. pombe types that are sensitive to chemicals. This helps them see if fission yeast can be used to study new medicines. For example, Doxorubicin is a common cancer medicine with many side effects. Scientists are using fission yeast to understand how Doxorubicin works. They are looking at genes linked to resistance. They found links between Doxorubicin's side effects and chromosome processes and how things move across cell membranes. Scientists are now using models of metabolism to find new drug targets. They expect more progress using fission yeast in the future.
How Scientists Study Fission Yeast
Fission yeast is easy to get, grow, and change to make new types. It can also be kept in either a haploid (single set of chromosomes) or diploid (double set of chromosomes) state. S. pombe is usually haploid. But when it's under stress, like not enough nitrogen, two cells will join. They form a diploid cell that then makes four spores inside a sac. This process is easy to see under a microscope. It allows scientists to study meiosis in a simpler way.
Almost any genetics experiment can be done with fission yeast. This includes:
- Tetrad dissection: Separating the four spores.
- Mutagen analysis: Studying changes in genes.
- Transformations: Adding new DNA to cells.
- Microscopy techniques: Like FRAP and FRET, which help see molecules in living cells.
New models, like Tug-Of-War (gTOW), are also used to study how strong yeast is and how genes are used. It is also quite easy to add or remove genes from fission yeast. Since its genome is fully sequenced, this task is well-known and easy to do.
See also
 In Spanish: Schizosaccharomyces pombe para niños
In Spanish: Schizosaccharomyces pombe para niños
