
Lens (vertebrate anatomy) facts for kids
Quick facts for kids Lens |
|
|---|---|
 |
|
| Lens of eye changing shape to focus near and far. | |
 |
|
| Schematic diagram of the human eye. | |
| Latin | lens crystallin |
| System | Visual system |
The lens (also called the crystalline lens) is a clear, curved part inside most land animal eyes. It works with the cornea and other clear fluids in your eye to bend light. This helps to focus images sharply onto the retina at the back of your eye.
In many animals, like humans, the lens can change its shape. This change helps the eye focus on objects that are close or far away. This amazing ability is called accommodation. It's a bit like how a camera lens adjusts to get a clear picture. Fish and other water animals often focus by moving their lens, not by changing its shape.
For humans, the lens is flatter at the front than at the back. When you are young, your lens is very powerful at focusing light. It provides about one-third of your eye's total focusing power. But as people get older, the lens becomes less flexible. This makes it harder to focus on close-up things. This is why many adults need glasses for reading.
Structure of the Eye Lens
Where the Lens Sits in Your Eye
The lens is located at the front of your eye. It sits behind the cornea (the clear outer layer) and the iris (the colored part). Tiny strings called suspensory ligaments hold the lens in place. These ligaments connect the lens to a muscle called the ciliary body.
Behind the lens is a jelly-like substance called the vitreous body. In front of the lens is a watery liquid called the aqueous humor. This liquid brings nutrients to the lens. In humans, the lens is shaped like a flattened sphere. An adult human lens is usually about 10 millimeters wide and 4 millimeters thick. It changes shape when you focus and grows slightly throughout your life.
Parts of the Lens




The lens has three main parts:
- The lens capsule
- The lens epithelium
- The lens fibers
The lens capsule is the tough, clear outer layer. Inside, the lens fibers make up most of the lens. The lens epithelium is a thin layer of cells between the capsule and the fibers at the front. The lens does not have nerves or blood vessels.
Scientists often describe the lens like a globe. The front and back are called the "poles." The outer edge is called the "equator." This equator area is where new cells are made. It is usually hidden by the iris.
The Lens Capsule


The lens capsule is a smooth, clear layer that completely covers the lens. It is very elastic, meaning it can stretch and bounce back. This elasticity helps the lens change shape when you focus. The capsule is made mostly of a protein called collagen. It is thickest near the equator and thinnest at the front and back poles. Tiny ligaments attach firmly to the capsule to hold the lens.
The Lens Epithelium
The lens epithelium is a single layer of cells at the front of the lens. These cells are very important for keeping the lens healthy. They bring nutrients into the lens and remove waste. They also help control the amount of water inside the lens.
A special thing about these cells is that they divide and create new lens fibers. This happens at the lens equator. The lens keeps adding new fibers from when you are a baby until you are old.
The Lens Fibers
The lens fibers make up most of the lens. They are long, thin, and clear cells. In humans, they can be up to 12 millimeters long. They are packed very tightly together, like the layers of an onion. These layers are called laminae.
The lens is divided into regions based on the age of its fibers. The oldest fibers are in the center, forming the "nucleus." Newer fibers are added to the outer layers, called the "cortex." As lens fibers mature, they lose their nuclei and other internal parts. This helps the lens stay clear.
Cell Structures and Spaces


New ways of looking at the lens show that it has small empty spaces and fluid-filled sacs. Scientists think these spaces help move nutrients and water through the lens. Some studies also suggest that lens cells can fuse together. This creates a network that helps with transport inside the lens.
How the Lens Develops

The development of the lens starts very early, when a human embryo is only about 4 millimeters long. Unlike most of the eye, which comes from deeper layers, the lens forms from the outer skin layer of the embryo.
First, a small group of cells from the inner embryo comes close to the outer skin. This makes the nearby skin start to change, forming a "lens placode." This placode is the first step in making the lens. It is a single layer of cells.
As the embryo grows, the lens placode deepens and curves inward. It eventually pinches off from the skin, forming a ball of cells called the "lens vesicle." This happens when the embryo is about 10 millimeters long.
Signals from the developing retina (the light-sensing part of the eye) then tell the cells at the back of the lens vesicle to grow longer. These signals also cause the cells to make special proteins called crystallins. Crystallins are clear proteins that help the lens bend light. These long cells fill the center of the vesicle, becoming the "primary fibers" of the lens. These primary fibers form the core of the adult lens.

More fibers, called "secondary fibers," are added from the lens epithelial cells at the equator. These new fibers grow towards the front and back, wrapping around the older fibers. As the lens gets bigger, the ends of these new fibers don't quite reach the poles. Instead, they form tight seams with neighboring fibers. These seams are called "sutures." The patterns of these sutures become more complex as more layers are added.
The lens continues to grow even after birth. New secondary fibers are added as outer layers. These new fibers are made by cells at the lens equator. As they mature, they also lose their nuclei and become clear lens fibers. In humans, the lens becomes more oval-shaped as it grows into early adulthood. After about age 20, it starts to become rounder again.
Many proteins control how the lens develops. One important gene is PAX6, which is like a master switch for lens formation. The lens is very useful for scientists studying how cells change and specialize. This is because older cells are pushed to the center and stay there, creating a record of development.
Different Lenses in Different Animals

In many water animals, like fish, the lens is much thicker and almost perfectly round. This helps them bend light more because water and the cornea bend light less than air and the cornea. Fish lens cells are also thinner than those in land animals.
Some animals, like diving birds, need to see well both above and below water. They have special lenses and corneas that let them change focus a lot. This helps them see clearly in both air and water. Human eyes, however, are not very good at seeing underwater.
How the Lens Works
Focusing Light


In humans, the main way the lens focuses is called accommodation. A well-known idea about how this works is the Helmholtz model. It describes how the lens changes shape.
External Forces on the Lens
The Helmholtz model says that the lens is normally held flat by tiny ligaments. These ligaments are pulled tight by the pressure inside your eyeball. When you want to focus on something close, a muscle called the ciliary muscle contracts. This relaxes the tension on the ligaments. Because the lens is elastic, it can then become rounder. This rounder shape increases its focusing power, letting you see close objects clearly.
To focus on something far away, the ciliary muscle relaxes. This makes the ligaments pull tighter on the lens again. The lens becomes flatter and thinner, which reduces its focusing power. This helps you see distant objects clearly.
Scientists are still studying exactly how the lens changes shape. There are other ideas, too, that suggest the ligaments and muscles work in more complex ways to fine-tune the lens's shape.
Internal Forces in the Lens
When the lens changes shape, it's not just a passive process. The lens itself might play an active role. For example, special proteins called aquaporins allow water to flow in and out of lens cells. Other proteins called connexins allow cells to communicate electrically.
The lens also has different layers with slightly different densities. This means the way light bends changes gradually from the center to the outside of the lens. This "gradient-index optics" helps the lens focus light even better. Scientists are learning that water movement inside the lens might help it change its overall focusing power.
Lenses in Birds, Reptiles, Fish, and Amphibians

In reptiles and birds, the ciliary body not only supports the lens but also has pads that press on the lens. These pads squeeze and release the lens to change its shape for focusing. This is different from mammals, where the suspensory ligaments usually do this job.
For vision in fish and amphibians, the lens usually stays the same shape. Instead, they focus by moving the lens forward or backward inside the eye. They use a muscle called the retractor lentis to do this.
In cartilaginous fish (like sharks), a small muscle pulls the lens forward to focus on nearby objects. In teleosts (bony fish), a muscle pulls the lens backward to focus on distant objects. Even though frogs and cartilaginous fish both move their lenses forward, the muscles they use are different.
The simplest vertebrates, like lampreys, don't have their lens attached to the eyeball. They flatten their cornea with outside muscles to push the lens backward and focus.
It's interesting to see how different animals, like vertebrates and mollusks, have developed similar eyes. Even though their eyes look alike on the outside, they use different types of cells and structures to achieve the same goal of focusing light.
Crystallins and Lens Clarity

Crystallins are special proteins that make up over 90% of the protein in your lens. These proteins are very good at packing tightly together. This tight packing helps the lens stay clear and increases its ability to bend light.
Another reason the lens is so clear is that mature lens fibers do not have parts that scatter light, like the nucleus or mitochondria. The lens fibers also have a strong internal "skeleton" that keeps them in their precise shape. If this skeleton is damaged, the lens can become cloudy.
The lens also blocks most ultraviolet (UV) light between 300 and 400 nanometers. Shorter UV wavelengths are blocked by the cornea. This is important because strong UV light can harm the retina. Artificial lenses used in eye surgery are also made to block UV light. People who don't have a lens (a condition called aphakia) can sometimes see UV light as a whitish-blue color.
How the Lens Gets Nutrients
The lens needs food to grow and stay clear. It doesn't need as much energy as other parts of the eye, though.
When a baby is developing, the lens is fed by a network of blood vessels. But these vessels disappear before birth. After birth, the lens gets all its food from the aqueous humor, the clear fluid at the front of the eye.

Nutrients flow into the lens from the front and back. Waste products flow out through the sides. This constant flow is maintained by special pumps in the lens cells. These pumps work with water channels and other molecules to move nutrients throughout the lens.
Glucose (a type of sugar) is the main energy source for the lens. Since mature lens fibers don't have mitochondria (the "powerhouses" of cells), they get most of their energy without using oxygen. This means the lens uses very little oxygen.
Lens Health Issues
- Cataracts are cloudy areas in the lens. Some are small and don't cause problems. Others can block light and make vision blurry. Cataracts often develop as people get older, but they can also happen from injuries or be present at birth. Diabetes can also increase the risk of cataracts. Cataract surgery involves removing the cloudy lens and replacing it with an artificial one.
- Presbyopia is when your eye loses its ability to focus on nearby objects as you get older. The exact reason is not fully known. But it's linked to changes in how hard, how shaped, and how big the lens becomes with age.
- Ectopia lentis means the lens has moved out of its normal position.
- Aphakia is when the lens is missing from the eye. This can happen after surgery, an injury, or sometimes a person is born without a lens.
Images for kids
-
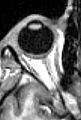
MRI scan of human eye showing lens.
-
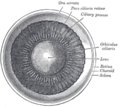
Inside the front part of the eye.
-

The crystalline lens, shown hardened and divided.
-

A magnified view of the lens edge, showing where epithelium turns into lens fibers.
-
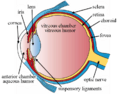
The structures of the eye labeled.
-
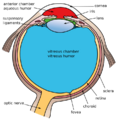
Another view of the eye with its structures labeled.
-

A diagram showing light rays, diaphragm, and crystalline lens.






See also
 In Spanish: Cristalino para niños
In Spanish: Cristalino para niños
- Accommodation reflex
- Crystallin
- Evolution of the eye, how the eye lens developed over time
- Intraocular lenses, artificial lenses
- Iris, the colored part of the eye
- Lens capsule
- Phacoemulsification, a type of cataract surgery
- Visual perception, how we see
- Zonules of Zinn, tiny fibers holding the lens
