
Reptile facts for kids
Quick facts for kids Reptiles |
|
|---|---|
 |
|
| Clockwise from above left: Green sea turtle (Chelonia mydas), Tuatara (Sphenodon punctatus), Nile crocodile (Crocodylus niloticus), and Sinai agama (Pseudotrapelus sinaitus). | |
| Scientific classification | |
| Extant groups | |
See text for extinct groups. |
|
 |
|
| Global reptile distribution (excluding birds) |

Reptiles are a group of animals with backbones that mostly live on land. The word "reptile" comes from Latin and means "one who creeps."
All living reptiles are cold-blooded. This means their body temperature changes with the temperature around them. They have scaly skin and lay eggs with tough shells. These eggs are called cleidoic eggs. Reptiles also get rid of waste as uric acid and have a cloaca. A cloaca is a single opening for waste and reproduction. Their heart and blood vessels are also different from mammals.
Many important reptile groups are now extinct. This includes the huge marine reptiles like ichthyosaurs and plesiosaurs. Dinosaurs were also reptiles, and birds are their living relatives. Today, the main surviving ancient reptiles are turtles, crocodiles, and the Tuatara. Most modern reptiles are snakes and lizards.
The study of living reptiles is called herpetology.
Contents
Reptiles and Birds
Some reptiles are more closely related to birds than to other reptiles. For example, crocodiles are closer relatives to birds than they are to lizards. Theropod dinosaurs are even closer relatives, because birds actually evolved from them.
Scientists who study cladistics group animals by their shared ancestors. This means they often include birds (over 10,000 species) with what we usually call reptiles. This larger group is known as Sauropsida.
Body Systems of Reptiles
Circulatory System

Most reptiles have a three-chambered heart. This heart has two upper chambers (atria) and one lower chamber (ventricle). They also have two main arteries (aortas) that send blood to the body. In these hearts, oxygen-rich and oxygen-poor blood can mix.
Some reptiles have special hearts. Crocodilians have a four-chambered heart, like mammals and birds. However, they can still control blood flow to bypass their lungs. Some snakes and lizards, like pythons, have three-chambered hearts that work like four-chambered hearts when they contract. This allows them to have strong blood pressure, similar to mammals.
Respiratory System
How Reptiles Breathe
All reptiles breathe using lungs. Some aquatic turtles can also absorb a little oxygen through their skin or cloaca. But they always need their lungs to breathe fully.
Different reptile groups breathe in different ways. Lizards and snakes mostly use their body muscles to breathe. These are the same muscles they use to move. This means they often have to hold their breath when they run fast. Some lizards, like monitor lizards, can also "buccal pump." This means they gulp air to help fill their lungs, allowing them to stay active longer.
Tegu lizards have a special muscle that helps their lungs expand more. Crocodilians have a muscle similar to our diaphragm. This muscle pulls a bone in their pelvis, which moves their liver. This creates space for their lungs to expand, helping them breathe.
How Turtles Breathe

It's tricky for turtles and tortoises to breathe because their shells are stiff. They can't expand their chests like other animals. Turtles have found many ways to solve this. Some have a muscle sheet around their lungs that helps them exhale. When they pull their limbs into their shell, they force air out. When they push their limbs out, they suck air in.
Turtles also use special muscles to move their internal organs up and down. This helps them breathe effectively. Some turtles can even breathe while walking or when completely inside their shells.
Reptile Palate
Most reptiles do not have a secondary palate. This means they have to hold their breath when they swallow food. Crocodilians have a bony secondary palate. This allows them to breathe even when they are underwater or eating. It also protects their brain from struggling prey.
Some skinks also have a bony secondary palate. Snakes have a different solution. Their windpipe (trachea) can stick out like a straw. This lets them breathe while swallowing large prey.
Skin

Reptile skin is covered in a tough, waterproof outer layer. This helps them live on dry land, unlike amphibians. Reptile skin is thinner than mammal skin.
The exposed parts of reptiles are protected by scales or scutes. These can sometimes have a bony base, forming armor. Lizards and snakes have overlapping scales all over their skin. These scales are part of their outer skin layer.
The scales on turtles and crocodiles are different. They are called scutes and grow from a deeper skin layer. In turtles, their body is hidden inside a hard shell made of fused scutes.
Excretory System
Reptiles mainly use two small kidneys to get rid of waste. Most reptiles that are diapsids (a group of reptiles) produce uric acid as their main waste. Turtles, like mammals, mostly excrete urea.
Reptile kidneys cannot make urine that is more concentrated than their body fluids. This is because they lack a special structure called a loop of Henle. Many reptiles use their colon to reabsorb water. Some can also use water stored in their bladder. Some reptiles also have special glands in their nose or tongue to get rid of extra salt.
Nervous System and Intelligence
The reptile nervous system is similar to an amphibian's. However, the reptile cerebrum and cerebellum (parts of the brain) are a bit larger. Most of their sense organs are well developed. Snakes do not have outer ears, but they do have inner ears. Reptiles have twelve pairs of cranial nerves.
Reptiles are generally thought to be less intelligent than mammals and birds. Their brain size compared to their body size is much smaller than in mammals. However, crocodiles have larger brains for their size and show complex social behaviors. Large lizards like monitors also show complex actions, including working together. Komodo dragons have even been seen playing.
Vision
Most reptiles are active during the day. Their vision is good for daylight, with color vision and good depth perception. Some snakes, like blind snakes, have very poor vision.
Some snakes have special heat-sensing pits. These pits are sensitive to infrared radiation (heat). They are very well developed in pit vipers, and also found in boas and pythons. These pits help snakes find warm-blooded prey, like rodents, in the dark.
Reproduction and Life Cycle


Most reptiles reproduce sexually. All reproductive activity happens through the cloaca. This is the single opening at the base of the tail for waste and reproduction.
Most reptiles lay amniotic eggs covered with leathery or hard shells. The developing baby (embryo) is protected inside the egg. There are no larval stages, meaning the young hatch looking like miniature adults.
Some lizards and snakes give birth to live young. This is called viviparity or ovoviviparity. Some species keep the eggs inside until they are ready to hatch. Others provide food to the developing young, similar to how mammals do with a placenta.
Some reptiles can reproduce without a mate. This is called parthenogenesis. It has been seen in some lizards, like geckos, and even Komodo dragons in captivity.
In some reptiles, the temperature of the eggs determines if they hatch as male or female. This is called temperature-dependent sex determination. It is common in turtles and crocodiles, and also happens in some lizards and tuataras.
Reptile Diet

Most reptiles are carnivorous (meat-eaters). They have simple and short guts because meat is easy to digest. Digestion is slower than in mammals. This is because reptiles are cold-blooded and have a lower metabolism. Their energy needs are much less than a mammal of the same size. Large reptiles, like crocodiles, can live for months on one big meal.
In the past, some very large reptiles were plant-eaters. Today, turtles are the main group of plant-eating reptiles. Some agams and iguanas also eat plants.
Plant-eating reptiles often swallow rocks or pebbles. These are called gastroliths. The rocks help grind up plant matter in their stomach. Sea turtles, crocodiles, and marine iguanas also use gastroliths as ballast. This helps them dive underwater.
Defense Mechanisms
Many small reptiles, like snakes and lizards, are prey for other animals. So, avoiding danger is their most common defense. When they sense danger, most snakes and lizards will crawl away. Turtles and crocodiles will dive into water and hide.

Reptiles also use camouflage to avoid being seen. They can blend into their surroundings using colors like gray, green, and brown.
If they can't escape, some reptiles will hiss loudly. Rattlesnakes shake their tail, which has hollow beads that make a rattling sound.
If these warnings don't work, reptiles use other defenses. Snakes have complex behaviors when attacked. Some will lift their head and spread their neck skin to look bigger. If that fails, some, like cobras and vipers, use venom. The venom is a changed saliva, delivered through fangs.
When a crocodile feels threatened, it will open its mouth wide to show its teeth and yellow tongue. If that doesn't work, it might start hissing. Then, it might change its body posture to look more intimidating, even inflating its body. If absolutely necessary, it might attack. Some crocodiles bite, others use their heads to smash opponents, or rush at the threat.

Geckos, skinks, and other lizards can drop part of their tail if caught. This is called autotomy. The detached tail wiggles, distracting the predator while the lizard escapes. The lizard can then regrow its tail over several weeks. The new tail will have cartilage instead of bone and might be a different color.
Living Reptiles
- Reptiles (living groups only)
- Order Crocodilia (crocodiles, gavials, caimans, and alligators): 23 species
- Order Sphenodontia (Tuatara from New Zealand): 2 species
- Order Squamata (lizards, snakes, and amphisbaenids ("worm-lizards"): about 7,000 species.
- Order Testudines (turtles, tortoises, and terrapins): approximately 300 species



-

Chameleons, a lizard
-

Komodo dragon, sleeping. Largest living lizard



How many species of reptiles are there?
There are about 10,000 species of traditional reptiles, and if we include birds, that number doubles! Lizards and snakes (squamates) today make up the vast majority of living reptiles.
| Reptile group | Described species | Percent of reptile species |
|---|---|---|
| Squamates | 9193 | 96.3% |
| - Lizards | 5634 | 59% |
| - Snakes | 3378 | 35% |
| - Amphisbaenians | 181 | 2% |
| Turtles | 327 | 3.4% |
| Crocodilians | 25 | 0.3% |
| Rhynchocephalians | 1 | 0.01% |
| Total | 9546 | 100% |
Images for kids
-

Reptiles, from Nouveau Larousse Illustré, 1897–1904. Notice that amphibians are included below the crocodiles.
-

"Antediluvian monster", a Mosasaurus discovered in a Maastricht limestone quarry, 1770 (contemporary engraving)
-

Modern classifications group "mammal-like reptiles", like this Varanodon, with other synapsids, not with living reptiles.
-

Bearded dragon (pogona) skeleton on display at the Museum of Osteology.
-

An early reptile Hylonomus.
-

A scene from the Mesozoic Era showing typical large reptiles: dinosaurs including Europasaurus holgeri, iguanodonts, and Archaeopteryx lithographica perched on the foreground tree stump.
-

A = Anapsid skull, B = Synapsid skull, C = Diapsid skull.
-

Varanus priscus was a giant meat-eating goanna lizard. It could be up to 7 meters long and weigh up to 1940 kilograms.
-

Skeleton of Champsosaurus, a choristodere. This was the last surviving group of extinct reptiles, living until about 11.3 million years ago.
-
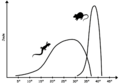
Energy output of a typical reptile versus a similar size mammal. Mammals have higher peak energy but only work in a small temperature range.
-

Skin of a sand lizard, showing the iconic scales of squamate reptiles.
-

A colubrid snake, Dolichophis jugularis, eating a legless lizard, Pseudopus apodus. Most reptiles eat meat, often other reptiles or small mammals.
-

Gastroliths (stomach stones) found from a plesiosaur.
-

The 1897 painting of fighting "Laelaps" (now Dryptosaurus) by Charles R. Knight.














See also
 In Spanish: Reptiles para niños
In Spanish: Reptiles para niños
