
Tasmaniosaurus facts for kids
Quick facts for kids Tasmaniosaurus |
|
|---|---|
 |
|
| Restored skull | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Archosauromorpha |
| Clade: | Crocopoda |
| Genus: | †Tasmaniosaurus Camp & Banks, 1978 |
| Type species | |
| †Tasmaniosaurus triassicus Camp & Banks, 1978
|
|
Tasmaniosaurus was an ancient reptile that lived in Tasmania, Australia, a very long time ago. Its name means "lizard from Tasmania," but it wasn't a true lizard. This animal lived during the Triassic period, about 250 million years ago.
Tasmaniosaurus is special because its fossil is one of the most complete ancient reptile skeletons found in Australia. It's also important because it was a very close relative of a group called Archosauriformes. This group includes dinosaurs, birds, and crocodiles.
Scientists first thought Tasmaniosaurus was part of a group called proterosuchids. But now, they believe it was a step between older reptiles and the first archosauriforms. Features like a hole in the skull near the eye, called an antorbital fenestra, and saw-like teeth were found in Tasmaniosaurus. This shows these features appeared earlier than once thought.
Contents
Discovering Tasmaniosaurus

The first mention of Tasmaniosaurus was in 1974. It was officially described by scientists Charles Lewis Camp and Maxwell Banks in 1978. They thought it was a type of archosaur, a group that includes dinosaurs and crocodiles. Another scientist, Tony Thulborn, agreed with this idea in 1986.
Over time, scientists changed how they defined "archosaur." Now, archosaurs only include animals that are closer to birds or crocodiles. Since proterosuchids, like Tasmaniosaurus, evolved before birds and crocodiles split, they are not considered true archosaurs anymore.
To fix this, scientists created a new group called Archosauriformes. This group includes proterosuchids and archosaurs. Archosauriformes is part of an even bigger group called Archosauromorpha. This group includes all reptiles more closely related to archosaurs than to lizards or snakes.
For a while, Tasmaniosaurus was not studied much. But in 2014, scientist Martin Ezcurra gave it a detailed new description. In 2016, he studied many archosauromorphs and found that the old group "proterosuchidae" was not correct. His study showed that Tasmaniosaurus was not a proterosuchid. Instead, it was the closest known relative to Archosauriformes, but not quite part of that group itself.
What Tasmaniosaurus Looked Like
Scientists know about Tasmaniosaurus from one main fossil skeleton. It's called UTGD 54655. This fossil includes parts of the skull, backbone, ribs, and leg bones. The skeleton is mixed up, and some parts are missing. But it's still one of the most complete reptile fossils from the Triassic period found in Australia.
Skull and Teeth
The skull of Tasmaniosaurus had some interesting features.
Snout Bones
The front part of its snout, called the premaxilla, was thought to be short at first. But later, scientists found it was similar in size to other archosauriforms. It was rounded at the front. Tasmaniosaurus likely had 6 or 7 teeth in each premaxilla.
The maxilla, a bone on the side of the snout, had a long row of teeth. Its shape suggests Tasmaniosaurus had an antorbital fenestra. This is a hole in the side of the snout, which is a key feature of archosauriforms. This hole shows how closely related Tasmaniosaurus was to archosauriforms. Each maxilla probably had about 21 teeth.
The lacrimal bone, located in front of the eye, was L-shaped. It looked very similar to the lacrimal bone of Proterosuchus. A bone from the roof of the mouth, called the pterygoid, was also found. It had six or seven teeth and looked almost the same as in Proterosuchus and Prolacerta.
Skull Roof
Parts of the skull roof were also found together. These included the frontals, parietals, and postfrontals. These bones are at the back of the skull. The frontals were long and not joined together. The parietals were also not joined and had wide, curved outer edges. These edges formed the inner part of large holes at the back of the head, called upper temporal fenestrae.
A large, crescent-shaped bone called the interparietal was at the very back of the skull roof. It was located between the parietal bones, similar to proterosuchids.
Lower Jaw
The main tooth-bearing bones of the lower jaw, called dentaries, were long, thin, and straight. This is like Prolacerta but different from most early archosauriforms, which had stronger or upward-curving jaws. The very tip of the jaw curved slightly downward. This made the first few teeth point a bit forward. Each dentary likely had more than 22 teeth.
Teeth
The teeth of Tasmaniosaurus were special. They were fused to the jaw bones and set in deep sockets. This is called ankylothecodont. Also, its teeth had serrated, saw-like edges. This is like archosauriforms but different from almost all other archosauromorphs. The teeth were curved and flattened from the side, making them knife-like. This suggests Tasmaniosaurus was a meat-eater.
Spine and Ribs
Vertebrae
The fossil has two vertebrae from the main part of the body. One was likely from near the neck, and the other from the middle of the back. These bones had tall neural arches and spines. The fossil also includes several tail vertebrae. The ones from the front of the tail were simpler than the back vertebrae. Those from the very end of the tail were very simple, without clear spines.
Ribs and Belly Ribs
Several ribs were found with the fossil. Some were curved. Tasmaniosaurus also had gastralia, which are like belly ribs. These bones got wider towards the back of the body, similar to Proterosuchus.
Other Bones
Shoulder Bone
The only part of the front limbs found was the interclavicle. This is a large, thin bone in the center of the shoulder area. The front part of this bone was wide and diamond-shaped. This is like some archosauromorphs, but different from most others, including Proterosuchus, which had a T-shaped interclavicle.
Leg and Foot Bones
The fossil also has several bones from the back legs. There are two tibia bones, which are shin bones. These were stronger than those of Proterosuchus. They had wide ends with a dip in the middle, similar to Prolacerta.
Both back feet were found, but they were mixed up and missing some bones. The outermost main foot bone, metatarsal V, had a hook-shaped end. Some toe bones and claws were also found. The claws were dull and only slightly curved.
How Tasmaniosaurus Lived
Brain and Senses

Scientists were able to study part of the Tasmaniosaurus' brain using a special mold called an endocast. This mold showed that its brain had large olfactory bulbs at the front. These parts of the brain are used for the sense of smell.
There were also wide, flat parts at the front of the brain. These might have been extra parts of the olfactory bulbs, meaning Tasmaniosaurus had an amazing sense of smell. If so, it suggests Tasmaniosaurus lived mostly on land, not in water. Animals that live in water usually have a weaker sense of smell.
Another idea is that these flat parts were a Jacobson's organ. This organ helps animals smell things by "tasting" the air. If Tasmaniosaurus had this organ, it would change what scientists thought about how this organ evolved in reptiles.
What Tasmaniosaurus Ate
The fossil of Tasmaniosaurus also had a bone from another animal. This bone was a maxilla, complete with teeth. Scientists first thought it belonged to an ancient amphibian. They believed it was food that Tasmaniosaurus had eaten.
However, later studies showed this was probably not true. The bone had too few teeth for an amphibian. It also had features that suggested it belonged to a small archosauriform, or even a young Tasmaniosaurus itself. So, it's unlikely this bone was a meal.
But, because Tasmaniosaurus had curved, saw-like teeth, scientists are almost certain it was a carnivore. This means it ate other animals.
Images for kids
-
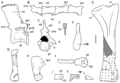
Illustrations of various vertebrae and limb bones of Tasmaniosaurus.
-

Skull roof bones of Tasmaniosaurus.
-

The interclavicle of Tasmaniosaurus (A) compared to that of Prolacerta (B) and Proterosuchus (C).



