
Baryonyx facts for kids
Quick facts for kids Baryonyx |
|
|---|---|
 |
|
| Baryonyx | |
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Superorder: | |
| Order: | |
| Suborder: | |
| Family: | |
| Subfamily: | |
| Genus: |
Baryonyx
Charig & Milner, 1986
|
| Species | |
|
B. walkeri Charig & Milner, 1986 (type) |
|

Baryonyx was a meat-eating dinosaur found in England and northern Spain. It lived about 125 million years ago, during the Early Cretaceous Period.
An amateur fossil hunter named William Walker first discovered it in 1983. Scientists, led by Alan J. Charig and Angela C. Milner, studied the skeleton. They named the dinosaur Baryonyx walkeri to honor its discoverer. Other good skeletons found later helped scientists learn a lot about this dinosaur.
Baryonyx could grow up to 8.5 meters (28 feet) long. Its huge claws were up to 35 centimeters (14 inches) long. This dinosaur likely ate fish, much like a modern grizzly bear does.
Contents
What Did Baryonyx Look Like?

In 2010, scientists estimated Baryonyx was about 7.5 meters (25 feet) long. It probably weighed around 1.2 metric tons (2,600 pounds). Earlier estimates in the 1990s suggested it might have been even larger. The first Baryonyx skeleton found was not fully grown. This means adult Baryonyx might have been bigger. Its relative, Spinosaurus, grew much larger, up to 15 meters (49 feet) long.
Its Head and Mouth
The skull of Baryonyx was long and narrow, like a crocodile's. It was about 95 centimeters (37 inches) long. The front part of its snout was rounded. Its nostrils were set far back on its head, not at the tip.
Baryonyx had a special bony roof in its mouth, similar to crocodiles. This helped it to hold onto prey. It also had a small ridge above its eyes. The bones around its eyes might have formed small horn-like bumps.
Baryonyx had many teeth, more than most other meat-eating dinosaurs. There were seven teeth in the front upper jaw. Most other theropods have only three to five. Its lower jaw had 32 teeth. The teeth were cone-shaped and slightly curved. They had fine serrations (like tiny saw teeth) on their edges. These teeth were perfect for gripping slippery fish. The teeth in the lower jaw were smaller and closer together than those in the upper jaw.
Its Body and Claws

Baryonyx's neck was shaped like a gentle "S". Its neck and back bones were lighter than they looked. They had hollow spaces, which helped reduce the dinosaur's weight.
Its shoulder blades were strong. The bones in its front limbs were short but very sturdy. The upper arm bone (humerus) was thick and wide. This suggests Baryonyx had powerful arm muscles.
The most famous part of Baryonyx was its huge thumb claw. This claw measured about 31 centimeters (12 inches) along its curve. In life, it would have been even longer with a keratin (like fingernail material) sheath. This large claw was sharp and slightly flattened. It likely helped Baryonyx catch and tear apart its food.
How Was Baryonyx Discovered?

On January 7, 1983, William J. Walker, a fossil enthusiast, found a large claw and some other bones. He found them in a clay pit near Ockley, England. A week later, he found the missing tip of the claw.
Scientists from the Natural History Museum, London then visited the site. They found many more bones. It took a team of eight museum staff and volunteers to dig out two tons of rock and dirt. Walker generously gave the claw to the museum. The company that owned the pit donated the rest of the skeleton. This area had been searched for 200 years, but no similar dinosaur remains had been found before.
Most of the bones were found inside hard rock lumps called nodules. Other Baryonyx fossils, mostly teeth, have since been found in other parts of the UK and Iberia (Spain and Portugal).
What Did Baryonyx Eat?

Scientists first thought Baryonyx was a fish-eater. They believed its long snout and many teeth were perfect for catching fish. They imagined it crouching by a riverbank, using its big claw to scoop fish out of the water. This is similar to how a grizzly bear fishes.
Some scientists disagreed, thinking it might have been a scavenger. They argued its jaws were too weak to kill large animals. However, a scavenger would still need strong jaws to break into carcasses.
Evidence of Its Diet

In 1997, scientists found direct proof of Baryonyx's diet. Inside the stomach area of the first skeleton, they found fish scales and teeth. These belonged to a common fish called Scheenstia mantelli. They also found bones from a young Iguanodon, another type of dinosaur. This showed that Baryonyx ate both fish and other dinosaurs. A possible gastrolith (stomach stone) was also found.
Scientists believe Baryonyx caught fish like a crocodile. It would grip fish with its long, narrow snout and then swallow them whole. It likely used its large claws to break up bigger fish or other prey. While it mainly ate fish, it was also an active hunter and would eat whatever it could find. It was not a top predator like Allosaurus.

Later studies supported the idea of Baryonyx being a fish-eater. Its skull structure was very similar to that of a gharial, a fish-eating crocodile. The special roof in its mouth helped its snout stay strong while catching prey.
In 2004, a spinosaurid tooth was found embedded in a pterosaur (flying reptile) bone. This proved that spinosaurids, including Baryonyx, did not eat only fish. They also hunted other animals.
Recent studies show that Baryonyx could widen its lower jaw. This allowed it to swallow large prey, like big fish or other animals. Its jaws were also similar to those of certain eels. This suggests they were good for grabbing prey in dark water.
How Did Baryonyx Move?

Scientists first thought Baryonyx might have walked on all fours sometimes. This would have been unique for a meat-eating dinosaur. However, later studies found no strong evidence for this. Its front limbs were strong, but probably not made for walking.
It's more likely that Baryonyx waded in water to hunt. Its strong front limbs and claws might have been useful for digging. It could have dug for water, excavated nests, or even dug for certain types of prey. Baryonyx probably did not fully submerge itself in water. It likely stayed in shallow areas, dipping its head in to catch fish.
Images for kids
-

Skeletal diagram of Baryonyx with the bones of Portuguese specimen ML1190 shown in red; it was originally considered a specimen of Baryonyx, but was since moved to Iberospinus
-

Reconstruction of the holotype skull, Museon, The Hague
-

Life restoration
-

Reconstructed skeleton in Japan
-

Skeletal diagram of the holotype specimen (below) compared with the related genus Suchomimus
-
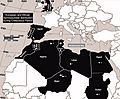
Distribution of spinosaurids in Europe and North Africa during the Cretaceous; 1, 3, 4, 5, 6 are Baryonyx
-

Restoration of Baryonyx with a fish
-
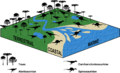
Spatial distribution of abelisaurids, carcharodontosaurids, and spinosaurids (the latter strongly associated with coastal environments)
-

Model carcass based on the position of the holotype bones, NHM







See also
 In Spanish: Baryonyx walkeri para niños
In Spanish: Baryonyx walkeri para niños
