
Euoplocephalus facts for kids
Quick facts for kids EuoplocephalusTemporal range: Upper Cretaceous 76.5–67 mya
|
|
|---|---|
 |
|
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Superorder: | |
| Order: | |
| Suborder: | |
| Infraorder: | |
| Family: | |
| Genus: |
Euoplocephalus
Lambe, 1910
|

Euoplocephalus (say "YOO-oh-PLO-sef-uh-lus") means "well-armored head." This big, plant-eating dinosaur was like a living tank! It was about 20 feet (6 meters) long and weighed around 2 tons. Its wide head had a tough, horn-like beak and small, peg-shaped teeth. It also had bony horns on its head. Euoplocephalus lived at the same time as fierce meat-eaters like Tyrannosaurus and Deinonychus.
Scientists have found fossils from over 40 different Euoplocephalus dinosaurs. These fossils were discovered in Alberta, Canada, and Montana in the United States. This makes Euoplocephalus the most well-known ankylosaurid dinosaur. Among the finds are 15 skulls, many teeth, and a few nearly complete skeletons with their armor still attached.
Contents
About Euoplocephalus
How Big Was It?

Euoplocephalus was one of the largest ankylosaurids. Only Ankylosaurus was bigger. This dinosaur was about 18 feet (5.5 meters) long and weighed around 2.5 tons. It was also very wide, about 8 feet (2.4 meters) across. Like other ankylosaurids, it had a broad, flat body that was low to the ground. It stood about four feet tall on its four short legs.
What Made It Special?
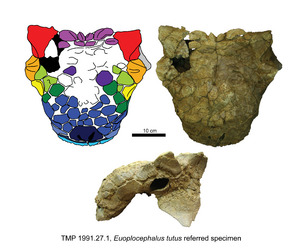
The skull of Euoplocephalus had some special features that help scientists tell it apart from other armored dinosaurs. For example, it had unique patterns of bony bumps on its face. It also had small bones above its eyes, which might have acted like bony eyelids. The front part of its nose was very shallow. Its upper jaw teeth curved inward, and the teeth themselves were small.
In 2013, scientists Victoria Arbour and Phil Currie pointed out other differences. For example, Euoplocephalus did not have round armor plates at the base of the horns on the back of its head. It also had large, rounded armor plates on the sides of its tail club.
Its Skeleton

The skull of Euoplocephalus looked like a triangle when seen from above. It was a bit wider than it was long. The largest skull found was about 16 inches (411 millimeters) long and 19 inches (478 millimeters) wide. Most of the bones in its skull were completely fused together. This meant that some openings usually found in dinosaur skulls had closed up.
Euoplocephalus had between 19 and 24 small teeth in each upper jaw. The very front of its snout, called the premaxilla, had no teeth. Its snout was blunt, wide, and tall. It had very complex air passages inside, which scientists are still trying to understand. Each side of its head had two nostrils. The lower jaw had 21 teeth.

Like most four-legged plant-eating dinosaurs, its neck was fairly short. Its shoulder blade was large and strong. Its front legs were very sturdy but shorter than its back legs. The tail was long and ended in a heavy, bony club.
Older drawings of Euoplocephalus sometimes show the tail club with two large spikes sticking up. This was a mistake based on an incomplete fossil. The spikes were actually located halfway along the tail, not at the very end. The end part of its tail was made stiff by bundles of hardened tendons. This helped it swing the tail club.
The backbone of Euoplocephalus had many fused bones. This made its back very strong. The hip bones were also very wide and flared out to the front. This helped support its large belly. Its thigh bones were short, strong, and straight. The shin bones were also robust and shorter than the thigh bones. Its feet were not well known but had three toes with hoof-like claws.
Its Armor

The head and body of Euoplocephalus were covered with bony armor. Only parts of its legs and the very end of its tail might have been unprotected. This armor was made of osteoderms, which are bony plates that grow in the skin. They are not part of the main skeleton.
Much of the armor was made of small, round bony pieces called ossicles. These were less than five millimeters across. Hundreds of these have been found with a single dinosaur. It is thought that many of these small pieces were fused together to form a kind of pavement. This pavement formed bands across the body, which allowed the dinosaur to move.
There were also rows of larger, oval-shaped armor plates mixed in with these bands. The largest and tallest plates might have been on its shoulders. Some of these large plates could be up to six inches (15 centimeters) long at their base. Large, keeled plates were also on its upper arms.
Its neck was protected by two bone rings, which were open on the underside. These are called "cervical half-rings." Each half-ring was made of six rectangular plates. Each plate had a large, keeled osteoderm on top.

The armor on its skull was made of many small bony plates called caputegulae ("head tiles"). These plates were fused with the skull bones. On the snout, they formed a messy, uneven pattern. On the back of the skull, the caputegulae fused into a single bony surface. The top edge of its eye sockets had two pyramid-shaped osteoderms. Euoplocephalus also had two pyramid-shaped "horns" growing from the back corners of its head. Below these, a large, tongue-shaped osteoderm stuck out from the lower rear side of the skull.
Life of Euoplocephalus
Movement and Social Life

Scientists believe Euoplocephalus could move as well as a modern rhinoceros or hippopotamus. Its strong upper arm muscles suggest its upper arm angled away from its body. Most Euoplocephalus fossils are found as single bones or partial skeletons. This might mean that these dinosaurs lived alone or in very small groups.
Protection and Senses
The armor of Euoplocephalus may have been covered in keratin, like our fingernails. Or it might have floated in the skin, similar to how armor works in modern crocodiles. Besides protecting the dinosaur, this armor had many blood vessels. This suggests it might have helped the dinosaur control its body temperature. The small bones above its eyes might have given extra protection to its eyes.
How It Defended Itself
The tail club of ankylosaurids is often thought of as a weapon for defense. In Euoplocephalus, only the end half of the tail was stiffened by hardened tendons. This allowed the front half of the tail to swing freely from side to side. The stiffened tendons would have helped transfer the force of the swing to the club.
A study in 2009 suggested that large ankylosaur tail clubs could hit hard enough to break bones. It is still unknown if they used their tails to fight other species, other Euoplocephalus, or both. The tail club could be swung low to hit the fragile leg bones of attacking meat-eating dinosaurs like theropods.
Euoplocephalus had relatively small eyes. However, this does not mean it had poor vision. The complex air passages in its skull suggest it had a very good sense of smell. Scientists also found an enlarged chamber at the back of its nose. This might have helped it smell even better.
A 2011 study found that the nasal passages of Euoplocephalus were looped and complex. This might have helped it control heat and water in its body. It also might have helped with making sounds. Researchers also rebuilt the dinosaur's inner ear. They found it could hear low-frequency sounds. They thought this might be an adaptation to hear low sounds made by its own nasal passages.
What It Ate
Like other ankylosaurians, Euoplocephalus was a herbivore, meaning it ate plants. It had a wide snout. This might mean it was not picky about what plants it ate, perhaps like a hippopotamus. This would have helped it find food without competing with other plant-eating dinosaurs that had narrower snouts.
Scientists first thought that ankylosaurians only moved their jaws up and down to eat. However, later research showed that Euoplocephalus could move its jaws forward and sideways. Its skull was also strong enough to handle a lot of force. This suggests Euoplocephalus could make more complex chewing movements. The way its teeth wore down and how its jaws connected shows that it likely pulled its lower jaws back and slightly inward while eating. This action would have helped it shear its food.
See also
 In Spanish: Euoplocephalus tutus para niños
In Spanish: Euoplocephalus tutus para niños
