
Cell signaling facts for kids
In biology, cell signaling (also called cell communication) is how cells talk to each other and to their surroundings. It's like a secret language that all living cells use, from tiny bacteria to complex animals. Cells need to receive, understand, and send signals to work properly.
These signals can come from outside the cell. They might be physical things like pressure, temperature, or light. Or, they can be chemical messages, like small molecules or special proteins. Cell signaling can happen over short distances, like between two cells right next to each other. It can also happen over long distances, like when hormones travel through your body.
Special proteins called receptors are key to cell signaling. They are like antennas that detect these chemical or physical signals. Receptors are usually found on the cell's surface or inside the cell, in places like the cytoplasm or nucleus. When a signal molecule (called a ligand) binds to a receptor, it causes a change in the receptor. This change then starts a chain reaction inside the cell.
This chain reaction is called signal transduction. It's like translating the signal into a chemical message the cell can understand. Sometimes, this directly opens a channel in the cell. Other times, it starts a "second messenger system." This system can make the signal much stronger, like turning a small whisper into a loud announcement.
Every cell is designed to respond to specific signals. This is super important for things like how you grow, how your body repairs itself, how your immune system fights germs, and how your body stays balanced (homeostasis). When there are mistakes in cell signaling, it can lead to diseases like cancer or diabetes.
Contents
How Cells Talk in Different Living Things
Cells talk to each other in many ways, depending on the organism.
Tiny Organisms
In small organisms like bacteria, cells use a cool trick called quorum sensing. Imagine a group of bacteria. They only start a big activity, like making light, when there are enough of them. They do this by sending out a signaling molecule. When many bacteria are present, there's a lot of this molecule, and they all "know" it's time to act together. This was first seen in a marine bacterium that makes light.
Slime moulds are another example. Individual slime mold cells come together to form a larger body and then spores. They are guided by a chemical signal, like a scent trail. They move towards where the chemical is strongest.
Plants and Animals
In plants and animals, cells signal in a few main ways:
- Short-distance signaling: Signals are released into the space around cells and affect nearby cells. This is called paracrine signaling.
- Long-distance signaling: Signals, like hormones, travel through the blood to reach distant cells. This is called endocrine signaling.
- Direct contact signaling: Cells touch each other to send signals. This is called juxtacrine signaling.
- Self-signaling: A cell releases a signal that then affects itself. This is called autocrine signaling.
How Signals Are Sent
Many cell signals are carried by molecules that one cell releases, and then they travel to another cell. These signaling molecules can be different types of chemicals, like fats, proteins, or even gases.
For example, in animals, special cells release hormones. These hormones travel through your blood to reach target cells in other parts of your body. These target cells then recognize and respond to the hormones. This is how your body controls many functions. Plants also use their own types of hormones to grow and respond to their environment.
Releasing Molecules: Exocytosis
One way cells release large amounts of molecules, like chemical messengers, is through a process called exocytosis. Think of it like a cell spitting out its contents. This process needs energy.
During exocytosis, small sacs called vesicles (which hold the molecules) move to the cell's outer membrane. They then join with the membrane and release their contents outside the cell. This is how brain cells (neurons) release neurotransmitters to send messages to other neurons.
Different Ways Cells Communicate
Cells communicate in various forms, each with a specific purpose.
Autocrine Signaling

In autocrine signaling, a cell sends out a chemical message, and then that same cell receives the message. It's like talking to yourself! This can cause changes within that cell.
Paracrine Signaling
Paracrine signaling is when a cell sends a signal to change nearby cells. The signaling molecules spread out over a short distance, affecting only cells in the local area. For example, neurotransmitters in your brain are paracrine signals. They travel across a tiny gap between neurons to send messages.
Some signaling molecules can act in different ways. For instance, epinephrine (also known as adrenaline) can be a hormone when released into your blood, making your heart race. But it can also be a neurotransmitter in your brain, sending messages between neurons.
Endocrine Signaling
Endocrine signaling uses signals called hormones. Hormones are made by special glands inside your body. They travel through your bloodstream to reach distant parts of your body. Only specific cells that have the right receptors will respond to a particular hormone. Your thyroid gland and adrenal glands are examples of endocrine glands.
Juxtacrine Signaling
Juxtacrine signaling happens when cells communicate through direct contact. This is common in multicellular organisms. There are a few ways this can happen:
- Two cells touch, and a molecule on one cell's surface connects with a protein on the other cell's surface.
- Cells are connected by special junctions that allow small molecules to pass directly between them.
- A cell's surface protein interacts with a protein in the extracellular matrix (the stuff around cells).
This type of signaling is important for your immune response and during development.
How Cells Receive Signals: Receptors

Cells receive information using special proteins called receptors. Receptors can bind to specific molecules (ligands) or react to physical things like light or pressure. When a target cell (a cell with the right receptor) detects a signal, the signal molecule binds to its receptor. This can happen on the cell surface or inside the cell.
When a signal molecule binds to a receptor, it often triggers a chain of events inside the cell. This can lead to various effects, like activating other molecules or changing how the cell behaves. For example, in humans, bacteria in your gut talk to each other and to your gut cells using signals.
Receptors on the Cell Surface
Cell surface receptors are super important for how cells work. If these proteins don't work right, it can lead to diseases like cancer or heart problems. These receptors are embedded in the cell's outer membrane. When a specific signal molecule binds to them, they change their shape. This change sends a message from outside the cell to the inside.
There are three main types of cell surface receptors:
- Ion channel linked receptors
- G protein–coupled receptors
- Enzyme-linked receptors
Ion Channel Linked Receptors

Imagine a gate in the cell membrane. Ion channel linked receptors are like these gates. When a chemical messenger (like a neurotransmitter) binds to them, the gate opens. This allows tiny charged particles called ions (like sodium or potassium) to flow into or out of the cell. This flow of ions changes the electrical charge of the cell.
For example, when a brain cell sends a message, it releases a neurotransmitter. This neurotransmitter binds to an ion channel linked receptor on the next brain cell. The channel opens, ions flow, and the message is passed along very quickly.
G Protein-Coupled Receptors

G protein–coupled receptors (GPCRs) are a very large group of receptors. They are called "seven-transmembrane receptors" because they weave through the cell membrane seven times. They detect molecules outside the cell and then start responses inside the cell by working with special proteins called G proteins.
GPCRs are found in all eukaryotes (organisms with complex cells), including animals and yeast. They can be activated by many different signals, like light, smells, hormones, and neurotransmitters. These receptors are involved in many diseases, which is why about half of all medicines today target GPCRs!
When a signal molecule binds to a GPCR, the GPCR changes shape. This change allows it to activate a G protein. The activated G protein then breaks into pieces, and these pieces go on to affect other proteins inside the cell, continuing the signal.
Enzyme-Linked Receptors

Enzyme-linked receptors are also on the cell surface. When a signal molecule binds to them, they don't just open a channel. Instead, they activate an enzyme part of themselves that is inside the cell. This enzyme then starts a chemical reaction.
Most enzyme-linked receptors are called "receptor tyrosine kinases." They add phosphate groups to other proteins, which is a common way to turn proteins on or off inside a cell.
Receptors Inside the Cell
Some receptors are not on the cell surface but are found inside the cell, in the cytosol or nucleus. These are called intracellular receptors.
Steroid Hormone Receptors
Steroid hormone receptors are a good example of intracellular receptors. Steroid hormones, like estrogen, are small and can easily pass through the cell's outer membrane. Once inside, they bind to their receptors. This binding causes the receptor to change. The receptor-hormone complex then often moves into the nucleus and affects which genes are turned on or off. This leads to changes in the cell that can take hours or even days to show up.
How Signals Travel Inside the Cell
When a signal molecule binds to a receptor, the receptor changes. This starts the process of signal transduction, which is like a relay race inside the cell. A series of different molecules, called relay molecules, pass the signal along.
This multi-step process has many benefits:
- Amplification: A small signal can be made much stronger. One activated receptor can lead to many relay molecules being activated.
- Regulation: There are many points where the signal can be controlled or adjusted.
- Fine-tuning: The cell can create a very specific response.
For example, a growth factor might bind to a receptor on the cell surface. This activates the receptor, which then adds phosphate groups to itself and other proteins. This creates new binding sites for other proteins, passing the signal along. One important pathway is the MAPK/ERK pathway. This pathway ultimately leads to changes in gene activity, which can make the cell grow and divide.
The final step of cell signaling is a specific cellular response. This can be almost any activity a cell does. It might be changing its shape, making a new protein, or even turning specific genes on or off in the nucleus. All these steps make sure that cells behave correctly, at the right time, and work together within the body.
In tiny organisms like bacteria, signaling can help them "wake up" from a resting state or work together to fight off viruses. In quorum sensing, many individual signals can create a strong group response. This might have even helped single-celled organisms evolve into multicellular ones!
Multicellular organisms also use signaling molecules called pheromones. These can warn others of danger, point to food, or help with reproduction.
Images for kids
-
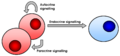
Different types of extracellular signaling
-

The AMPA receptor bound to a glutamate antagonist showing the amino terminal, ligand binding, and transmembrane domain, PDB 3KG2
-
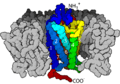
A G Protein-coupled receptor within the plasma membrane.
-
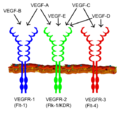
VEGF receptors are a type of enzyme-coupled receptors, specifically tyrosine kinase receptors
-

Key components of a signal transduction pathway (MAPK/ERK pathway shown)
-
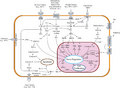
Signal transduction pathways that lead to a cellular response
-
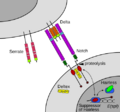
Notch-mediated juxtacrine signal between adjacent cells.



See also
 In Spanish: Comunicación celular para niños
In Spanish: Comunicación celular para niños
- Scaffold protein
- Biosemiotics
- Molecular cellular cognition
- Crosstalk (biology)
- Bacterial outer membrane vesicles
- Membrane vesicle trafficking
- Host-pathogen interface
- Retinoic acid
- JAK-STAT signaling pathway
- Imd pathway
- Localisation signal
- Oscillation
- Protein dynamics
- Systems biology
- Lipid signaling
- Redox signaling
- Signaling cascade
- Cell Signaling Technology, an antibody development and production company
- Netpath – A curated resource of signal transduction pathways in humans
- Synthetic Biology Open Language
- Nanoscale networking – leveraging biological signaling to construct ad hoc in vivo communication networks
- Soliton model in neuroscience—Physical communication via sound waves in membranes
- Temporal feedback
