
Edmontosaurus facts for kids
Quick facts for kids EdmontosaurusTemporal range: Upper Cretaceous 73–65.5 mya
|
|
|---|---|
 |
|
| Mounted skeleton of E. regalis, Oxford University Museum of Natural History | |
| Scientific classification | |
| Kingdom: | |
| Class: | |
| Superorder: | |
| Order: | |
| Suborder: | |
| Family: | |
| Genus: |
Edmontosaurus
|
Edmontosaurus was a huge, plant-eating, duck-billed dinosaur that lived during the very end of the Cretaceous period. It had short arms, a long, pointed tail, and feet with three toes that looked like hooves. Its hands were shaped like mittens. Its head was flat and sloped, with a wide, toothless beak. Inside its cheeks, it had hundreds of teeth packed closely together to grind its food.
Many Edmontosaurus fossils have been found in Alberta, Canada, and in Alaska, Wyoming, Montana, and New Jersey in the USA. Two amazing mummified Edmontosaurus fossils were even discovered in Wyoming, showing preserved skin!
Scientists think Edmontosaurus liked to live near coasts and flat coastal areas because of where its fossils are found. It could walk on two legs or on all four. Since many bones have been found together in "bone beds," it's believed that Edmontosaurus lived in large groups, like herds. They might have even traveled long distances, like migratory animals do today.
Contents
What did Edmontosaurus look like?

Edmontosaurus was a big, strong dinosaur, like other duck-billed dinosaurs (called hadrosaurids). It had a long, flat tail and a head that looked like a duck's beak. Unlike some other hadrosaurids, its skull didn't have a bony crest. Instead, it might have had a soft, comb-like crest made of skin and scales on its head.
Its front legs were not as strong as its back legs, but they were long enough for it to stand or move on all fours. Edmontosaurus was one of the largest hadrosaurids. A full-grown adult could be about 9 meters (30 feet) long, and some very large ones reached 12 to 13 meters (39 to 43 feet) long. They weighed around 4 metric tons (about 8,800 pounds), which is like two large cars!
How big was its skull?

The skull of a grown Edmontosaurus could be over a meter (3 feet) long. It was shaped like a triangle when viewed from the side. From above, the front of the skull was wide, forming a duck-bill shape. The beak itself had no teeth and was covered in a tough, horn-like material, similar to a bird's beak.
Edmontosaurus only had teeth in the back of its jaws, in its cheeks. These teeth were always growing and being replaced, taking about six months to form. They were arranged in columns, with up to six teeth in each column, forming a "dental battery." This allowed them to grind up tough plant material.
What about the rest of its body?

The number of bones in its spine (vertebrae) varied a bit between individuals. Edmontosaurus had a strong back and tail, stiffened by special ossified tendons. These tendons were like bony rods that crisscrossed along its spine, making its back and part of its tail very straight and strong. This helped support its large body.
Its shoulder blades were long and flat. The hips were made of three parts. The way its hips were shaped meant it couldn't stand straight up with its back perfectly vertical.
The front legs were shorter and less strong than the back legs. Its upper arm bone had a large area for muscles to attach. Its wrists were simple, with only two small bones. Each hand had four fingers, but no thumb. The second, third, and fourth fingers were about the same length and were covered in skin, making them look like a mitten. The little finger was much shorter and stuck out.
The thigh bone was thick and straight, with a strong ridge for powerful muscles. These muscles helped pull the legs backward and kept the tail balanced. Each foot had three toes with hoof-like tips.
Did it have skin?

Yes! Several Edmontosaurus annectens fossils have been found with amazing skin impressions. One famous fossil, nicknamed "Dakota," even had some of its original skin compounds preserved! These finds show us what its scales looked like over most of its body.
Some E. regalis fossils also show well-preserved skin, including one that suggests it had a soft crest or wattle (like a turkey's wattle) on its head. We don't know if E. annectens had this crest, or if it was different between male and female dinosaurs.
How was Edmontosaurus discovered?
The first well-known species of Edmontosaurus was first named Claosaurus annectens in 1892 by Othniel Charles Marsh. This was based on a partial skull and skeleton found in Wyoming. These fossils were important because they were among the first nearly complete dinosaur skeletons to be put on display in museums in the United States.
The name Edmontosaurus itself was given in 1917 by Lawrence Lambe. He named it after the Edmonton Formation (now called the Horseshoe Canyon Formation) in Alberta, Canada, where two partial skeletons were found. The type species (the main example of the genus) is E. regalis, meaning "regal" or "king-sized," because it was so large.
Over the years, scientists have given Edmontosaurus and its close relatives many different names. But thanks to new studies, we now know that many of these names were actually for the same two species, or even just different growth stages of those species!
What species of Edmontosaurus are there?

Today, scientists generally agree that there are two main species of Edmontosaurus:
- E. regalis: This species is found only in the Horseshoe Canyon Formation in Alberta, Canada. It lived during the late Campanian stage of the Late Cretaceous period.
- E. annectens: This species is found in several rock formations in Saskatchewan, Montana, South Dakota, and Wyoming. It lived during the later Maastrichtian stage of the Late Cretaceous.
Scientists used to think Anatotitan copei and E. saskatchewanensis were separate species, but now they believe these were just different growth stages of E. annectens. E. saskatchewanensis was likely a young E. annectens, and Anatotitan copei was a fully grown adult. This means that as Edmontosaurus grew, its skull changed shape quite a bit, becoming longer and flatter.
E. annectens generally had a longer, lower, and less bulky skull compared to E. regalis. Even though they look a bit different, all E. regalis fossils come from older rocks than E. annectens fossils.
How did Edmontosaurus live?
How did it grow?

Studies have shown that the skulls of Edmontosaurus changed a lot as the dinosaurs grew up. Many features that scientists once used to name new species were actually just signs of different growth stages. For example, a species once called Thespesius edmontoni is now thought to be a young E. regalis. This helps scientists understand how these dinosaurs developed from juveniles to adults.
What did it eat?

As a hadrosaurid, Edmontosaurus was a large plant-eater that lived on land. Its teeth were always being replaced and were packed into "dental batteries" with hundreds of teeth. Only a few were used at a time. It used its wide beak to cut off plants, perhaps by cropping them or by closing its jaws like a clamshell over twigs and branches to strip off leaves.
Scientists believe Edmontosaurus and other similar dinosaurs had cheek-like structures, either muscular or not. These cheeks would have helped keep food inside its mouth while it chewed. It could eat plants from the ground up to about 4 meters (13 feet) high.

Studies of the scratches on Edmontosaurus teeth show that it moved its jaws in a complex way, not just up and down. This motion helped it grind its food very effectively. The scratches suggest that Edmontosaurus was a grazer, meaning it ate plants close to the ground, like grass or low-growing ferns. It might have even accidentally eaten some dirt, which would have added to the scratches on its teeth.
How did it move?

Like other duck-billed dinosaurs, Edmontosaurus could walk on two legs or four. It probably moved on all fours when standing still or moving slowly. When it needed to go faster, it would switch to using only its powerful hind legs.
Computer models suggest that Edmontosaurus could run quite fast, possibly up to 45 kilometers per hour (28 miles per hour)! This means it was quick enough to escape from predators.
Did it fight with other dinosaurs?

Edmontosaurus lived at the same time and in the same places as Tyrannosaurus rex. An adult E. annectens skeleton on display in the Denver Museum of Nature and Science shows clear signs of a theropod bite on its tail. The damage to its tail bones suggests an attack from the right rear. The top of the tail was high enough that only a large theropod like T. rex could have made such a bite.
The bones show that the Edmontosaurus survived the attack, as the injuries had partially healed. This suggests it might have escaped or even used its strong tail to fight back against the Tyrannosaurus. Scientists also noticed an older, healed fracture in its hip, which might have made the Edmontosaurus limp and an easier target for the T. rex.
Images for kids
-

Skeletal drawing of the E. annectens (then Claosaurus) by Othniel Charles Marsh.
-

Older E. annectens skeleton displays, once called Anatosaurus.
-

Skin impression from the famous "Dakota" fossil, found in 1999.
-

Mounted skeletons of a young and adult E. annectens at the Houston Museum of Natural Science.
-
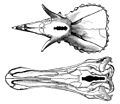
A 1905 chart showing the relatively small brains of a Triceratops (top) and E. annectens.
-

E. annectens skull showing the keratinous beak (partially removed on the right side), at the Natural History Museum of Los Angeles County.
-

Horseshoe Canyon Formation near Drumheller, where Edmontosaurus fossils were found. The dark bands are coal seams.
-

Edmontosaurus fossils found in eastern Montana (Hell Creek Formation).
-
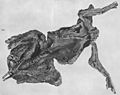
Possible stomach contents were reported from the "Trachodon mummy" at the American Museum of Natural History, but were never fully described.
-

The Edmontosaurus specimen at the Senckenberg Museum, showing preserved skin.
-

Edmontosaurus annectens displayed as being hunted by a Tyrannosaurus, showing how they might have interacted.











See also
 In Spanish: Edmontosaurus para niños
In Spanish: Edmontosaurus para niños
