
Red imported fire ant facts for kids
Quick facts for kids Red imported fire ant |
|
|---|---|
 |
|
| A group of fire ant workers | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Formicidae |
| Subfamily: | Myrmicinae |
| Genus: | Solenopsis |
| Species: |
S. invicta
|
| Binomial name | |
| Solenopsis invicta Buren, 1972
|
|
| Script error: The function "autoWithCaption" does not exist. | |
| Synonyms | |
|
|
Script error: No such module "Check for conflicting parameters".
The red imported fire ant (Solenopsis invicta) is also known as the fire ant or RIFA. This ant comes from South America. It belongs to a group of ants called Solenopsis. Scientists first described this ant in 1916. Later, in 1972, it was given the name invicta, which means "unconquered" or "invincible".
Contents
What's in a Name?
The name invicta comes from Latin and means "unconquered". It reminds us of the old phrase Roma invicta, which meant "unconquered Rome". The name for its group, Solenopsis, comes from Ancient Greek. It means "pipe appearance".
People call this ant the "red imported fire ant" because of its color and where it came from. The "fire ant" part is because its sting feels like a burning fire! In Brazil, some people call it toicinhera, which means "pork fat ant".
What Do Fire Ants Look Like?

Red imported fire ants are usually red or yellowish. Their back part, called the gaster, can be brown or black. Male fire ants are completely black.
Different Sizes of Workers

These ants are polymorphic. This means the worker ants in a colony come in different shapes and sizes.
Where Do Fire Ants Live?

Red imported fire ants like to live in many different places. You can find them in rainforests, deserts, and grasslands. They also live near roads, buildings, and even inside electrical equipment.
Their nests are large mounds made of soil. You usually won't see an entrance to the nest. This is because the ants build tunnels to find food, and they come out far away from their home.

Even though they are from South America, these ants have spread to many other places. They have been found in Australia, New Zealand, parts of Asia, the Caribbean, Europe, and the United States.
How Do Fire Ants Behave?
Fire ants have some interesting behaviors. For example, if water levels rise, they can build floating "rafts" with their bodies to survive! They also clean up their nests. They carry dead ants and trash to special piles outside their home. This is called necrophoric behavior.
Reproduction and Mating
Mating flights happen when the weather is warm. During these flights, male and female ants with wings (called alates) fly and mate. This can last for up to 30 minutes.
How Fire Ants Talk
Worker ants talk to each other using special chemicals. These chemicals, called pheromones, help them find food, recruit other ants, and defend their colony.
Ant Colony Life
A new colony can be started by one queen or a group of queens. If there are many queens, they will compete to be the main queen once the first worker ants appear. Worker ants live for several months, but queens can live for many years. A single colony can have anywhere from 100,000 to 250,000 ants!
There are two main types of fire ant colonies:
- Polygynous colonies: These nests have many queens.
- Monogynous colonies: These nests have only one queen.
What Do Fire Ants Eat?

Fire ants look for food on warm or hot days. They can also stay out at night. They are omnivores, meaning they eat both plants and animals. Their diet includes dead mammals, arthropods, insects, and seeds. They also like sweet things, like honeydew from other insects. They sometimes have special relationships with these insects.
Who Eats Fire Ants?
Some animals eat fire ants. These include arachnids (like spiders), birds, and many insects. Other ants, dragonflies, earwigs, and beetles are also predators of fire ants.
Ants and People
Fire Ant Stings
Venom is very important for fire ants. They use it to catch food and to defend themselves. About 95% of their venom is made of special chemicals called solenopsins. The rest is a mix of proteins that can be very strong for some people. The name "fire ant" comes from the burning feeling their sting causes.
Many people in the United States get stung by fire ants every year. Some people can become allergic to the venom. Most people who are stung feel a strong burning and swelling. Then, small bumps called pustules may form, which can stay for several days.
A small number of people might have a serious allergic reaction called anaphylaxis. This reaction can be very dangerous if not treated quickly. Symptoms can include dizziness, chest pain, feeling sick, sweating a lot, low blood pressure, and trouble breathing. If someone has a severe reaction, they need immediate medical help, often an injection of epinephrine.
Fire Ants as Pests
.jpg)
Fire ants are known as a big pest. They cause a lot of damage each year and can harm wildlife. They love living in cities, and their presence can make people not want to do outdoor activities.
Their nests can be built under things like sidewalks and building foundations. This can cause problems for structures or even make them fall apart. Besides damaging buildings, fire ants can also harm equipment and businesses. They can even lower the value of land and property.
In farming, fire ants can damage crops and farm machines. Mounds built in fields can make it hard to harvest crops. They can also hurt or even kill animals and livestock, especially young or sick ones.
However, fire ants can sometimes be helpful. They eat other insects that are pests on crops. To control these ants, people often use baits or fumigation. Because they are such a big problem, fire ants are one of the most studied insects on Earth!
Images for kids
-

Paratype specimen of S. invicta collected from Brazil
-

Casent label of S. invicta paratype worker
-

Whole body image of a worker
-

Electron micrograph of a hatching fire ant egg
-

Photomicrograph of sting apparatus components
-

Water loss rates of workers and female alates in S. invicta and S. richteri
-

A floating "raft" of red imported fire ants (RIFA) in North Carolina is seen over land that normally forms the bank of a pond. The land had become submerged due to excessive rain and resultant flooding which inundated the nest. The raft is anchored to some blades of grass extending above the water's surface.
-

Worker foraging on a common sunflower (Helianthus annuus)
-

L. mactans feeding on a captured insect: Red imported fire ants constitute 75% of all prey items captured by this spider.
-

Newly hatched phorid fly emerging from a host's head
-

Male alate
-
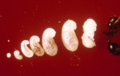
Lifecycle of the red imported fire ant, including several larval instars
-

Pupa of a queen
-

Predation of the Stock Island tree snail by red imported fire ants is believed to be a major cause of its extinction in the wild
-

The red imported fire ant and Phenacoccus solenopsis have formed a symbiotic relationship with each other
-

Mounds found on farmland
-

Bait used against the ant
-

Exposed red imported fire ant mound

















See also
 In Spanish: Hormiga roja de fuego para niños
In Spanish: Hormiga roja de fuego para niños
