
Sea urchin facts for kids
Quick facts for kids Sea urchin |
|
|---|---|
 |
|
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: |
Echinoidea
|
Sea urchins are amazing sea creatures that belong to a group called Echinodermata. This name means "spike-skinned," which makes sense because sea urchins are covered in spines! They live only in the ocean and are usually shaped like a globe. Their bodies are protected by strong plates made of calcium carbonate and those famous spines. The word "urchin" used to mean hedgehog, and in many languages, sea urchins are still called "sea hedgehogs."
Like other echinoderms, sea urchins have a special five-part symmetry (called pentamerism). This means their body parts are often arranged in fives around a central point. You might not notice this on a living sea urchin, but it's easy to see on their dried-out shell. They move around using hundreds of tiny, clear, sticky 'tube feet'.
Sea urchins mostly eat algae (seaweed) and small animals. They have a unique chewing tool called Aristotle's lantern. It's named after the Greek philosopher Aristotle because he was very interested in sea urchins. This tool helps them scrape off food stuck to surfaces as they move.
Contents
What are Sea Urchin Spines Like?
Sea urchin spines can be long and sharp in some types. These spines help protect the urchin from predators. If a spine pokes your skin, it can be painful, but it's usually not dangerous. Sea urchins also have tiny, pincer-like tools called pedicellariae all over their body, hidden between the spines. These little pincers help keep the urchin's shell clean by grabbing anything that tries to settle on it, like tiny larvae. We don't know if the spines themselves are venomous, but the pedicellariae can be.
Most sea urchins have spines that are about 1 to 3 centimeters long and 1 to 2 millimeters thick. They are not extremely sharp. However, a sea urchin called Diadema antillarum, found in the Caribbean, has thin, very long spines that can reach 10 to 30 centimeters! These can be quite dangerous.
Do Sea Urchins Have Teeth?
Yes, sea urchins have five jaws, and each jaw has one tooth. These teeth keep growing throughout the sea urchin's life. For example, the teeth of the pink sea urchin rub against each other, which helps them stay sharp.
What Do Sea Urchins Eat?

Sea urchins mainly eat algae, but they can also eat other sea creatures. This includes sea cucumbers and many different invertebrates like mussels, polychaete worms, sponges, and brittle stars.
Who Eats Sea Urchins?
Sea urchins are a favorite food for animals like sea otters and wolf eels. If there aren't enough predators to eat sea urchins, their numbers can grow very large. When this happens, they can eat almost all the nearby kelp (a type of large seaweed). This creates an urchin barren, which is an area of the ocean floor that is empty of large seaweeds and the animals that live among them. For example, when sea otters returned to British Columbia, they ate many sea urchins. This helped the coastal ecosystems become much healthier.
How Old Are Sea Urchins?
The oldest sea urchin fossils we've found are from the Lower Ordovician period. This was a very long time ago! The first sea urchins had flexible shells, where the plates could slide over each other. For a long time, sea urchins were not a very common part of ocean life. Only one group of sea urchins survived a huge extinction event to become the ancestors of all modern sea urchins. This group, called Cidaroids, then developed into all the other types we see today.
Around the Jurassic period, some sea urchins started to lose their perfect five-part symmetry. This meant they developed a clear front and back end. This change allowed them to live in new places, especially by burrowing into the sand. Sand dollars and heart urchins are examples of these successful burrowing sea urchins. Today, living sea urchins are an important part of ocean life, especially in shallow waters near the shore.
Do People Eat Sea Urchins?
Yes, some people eat the reproductive parts of sea urchins, which are often called "roe." This is very popular in Korea. In Japan, this type of sushi is called uni.
.jpg)
How Were Sea Urchins Used as Tools?
In the Pacific Ocean, people used bunches of heavy spines from slate-pencil urchins (Heterocentrotus species) as tools. They used them to rub and smooth surfaces when building ships.
Sea Urchins in Aquariums
.jpg)
Some types of sea urchins, like the slate pencil urchin (Eucidaris tribuloides), are often sold in aquarium stores. Certain species are good at controlling stringy algae in fish tanks. They make great additions to a tank that has other invertebrates.
Sea Urchins in Old Stories
In Denmark and southern England, there was an old folk tale about sea urchin fossils. People thought they were "thunderbolts" and could protect against lightning or witchcraft. They were seen as a lucky charm. Another story said they were petrified (turned to stone) snake eggs. These "snake eggs" were believed to protect against heart and liver diseases, poisons, and injuries in battle. So, people carried them as amulets. The legend said these "eggs" were magically created from foam made by snakes at midsummer.
Images for kids
-
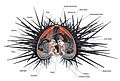
Sea urchin anatomy based on Arbacia sp.
-

Dentition of a sea urchin
-

Aristotle's lantern in a sea urchin, viewed in lateral section
-
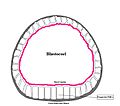
Sea urchin blastula
-

The development of a regular sea urchin
-

Pluteus larva has bilateral symmetry.
-

The flower urchin is a dangerous, potentially lethally venomous species.
-

The thick spines (radiola) of Cidaridae were used for walking on the soft seabed.
-

Paracentrotus lividus, a regular sea urchin (Euechinoidea, infraclass Carinacea)
-

A sand dollar, an irregular sea urchin (Irregularia)
-

Phyllacanthus imperialis, a cidaroid sea urchin (Cidaroidea)
-

Test of an Echinus esculentus, a regular sea urchin
-
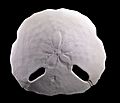
Test of an Echinodiscus tenuissimus, an irregular sea urchin ("sand dollar")
-

Test of a Phyllacanthus imperialis, a cidaroid sea urchin. These are characterised by their big tubercles, bearing large radiola.
-

Close-up of the test showing an ambulacral groove with its two rows of pore-pairs, between two interambulacra areas (green). The tubercles are non-perforated.
-

Wolf eel, a highly specialized predator of sea urchins
-

A sea otter feeding on a purple sea urchin.
-

A crab (Carpilius convexus) attacking a slate pencil sea urchin (Heterocentrotus mamillatus)
-

A wrasse finishing the remains of a damaged Tripneustes gratilla
-

Purple sea urchins at low tide in California. They dig a cavity in the rock to hide from predators during the day.
-

Dermechinus horridus, an abyssal species, at thousands of meters deep
-

Antarctic sea urchin (Sterechinus neumayeri) inhabits frozen seas.
-

The shape of the shingle urchin allows it to stay on wave-beaten cliffs.
-

Archaeocidaris brownwoodensis, Cidaroida, Carboniferous, c. 300 mya
-

Miocidaris coaeva, Cidaroida, Middle Triassic, c. 240 mya
-

Clypeus plotti, Irregularia, Middle Jurassic, c. 162 mya
-

Echinocorys, Holasteroida, Upper Cretaceous, c. 80 mya
-

Echinolampas ovalis, Cassiduloida, Middle Eocene, c. 40 mya
-

Clypeaster portentosus, Clypeasteroida, Miocene, c. 10 mya
-

Japanese uni-ikura don, sea urchin egg and salmon egg donburi
-

Open sea urchins in Sicily



.JPG)

_early_pluteus_width_ca.JPEG)

.JPG)






.jpg)

















See also
 In Spanish: Erizos de mar para niños
In Spanish: Erizos de mar para niños
