
Cardabiodon facts for kids
Quick facts for kids Cardabiodon |
|
|---|---|
 |
|
| Holotype and paratype teeth of Cardabiodon venator from the Fairport Member of the Carlile Shale in Mosby, Montana. | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Chondrichthyes |
| Order: | Lamniformes |
| Family: | †Cardabiodontidae |
| Genus: | †Cardabiodon Siverson, 1999 |
| Type species | |
| †Cardabiodon ricki Siverson, 1999
|
|
| Species | |
|
|
| Synonyms | |
|
|
Cardabiodon (meaning 'Cardabia tooth') was a huge mackerel shark that lived about 95 to 91 million years ago (Ma). It swam in the oceans during the Late Cretaceous period. This shark belonged to a special family called Cardabiodontidae. This family had unique teeth.
There are two known types (species) of Cardabiodon: C. ricki and C. venator. Scientists have found Cardabiodon fossils in places like Australia, North America, England, and Kazakhstan. It was likely a shark that lived in cooler, temperate waters, much like the modern porbeagle shark.
Cardabiodon was one of the biggest sharks of its time. It could grow up to 5.5 meters (18 feet) long. This shark was probably a top predator in its ocean home. It used its large, strong teeth and fast swimming skills to hunt other marine animals. Its prey included plesiosaurs, ichthyosaurs, and other big fish. Cardabiodon also had a very heavy and strong body.
Scientists have learned some things about its life. Baby Cardabiodon sharks were about 41 to 76 centimeters (16-30 inches) long when they were born. They became adults when they were around five to seven years old. However, we don't know how long Cardabiodon sharks lived in total.
Discovering Cardabiodon

How it was found
The first Cardabiodon fossils were found in Western Australia. Paleontologist Mikael Siverson described them in 1999. These fossils were from the Gearle Siltstone, a rock layer from about 96 to 94.7 million years ago. The first discovery included about 100 teeth and 15 backbones from a young shark. Later, more teeth were found from the same shark.
Siverson named this new shark Cardabiodon ricki. The name Cardabiodon means "Cardabia tooth." Cardabia is the place in Australia where it was found. The name ricki honors Rick French, who owned the land where the fossils were discovered.
In 2005, a second type of Cardabiodon was found in Montana, USA. This species was named C. venator. Its name means "hunter" in Latin. This name was chosen because scientists believe C. venator was a top hunter in its ancient ocean.
Shark family tree
Cardabiodon was a unique shark because of its special teeth. But it is still grouped with other mackerel sharks. This is because it shares some basic tooth and backbone features with them. To show how unique Cardabiodon was, Siverson created a new family for it: Cardabiodontidae.
Another Late Cretaceous shark, Dwardius, has teeth very similar to Cardabiodon. Scientists think these two sharks might be closely related. Some even wondered if they were the same type of shark. However, scientists have found differences in how their teeth changed over time. This suggests they are separate but related groups.
Scientists are still working to fully understand where the Cardabiodontidae family fits in the larger shark family tree. A tooth from C. ricki found in Alberta, Canada, showed features of both C. ricki and C. venator. This suggests that C. ricki might have slowly changed into C. venator over time.
Scientists have found fossils of even older sharks that are related to Cardabiodon. One notable find is from Australia, dating back 105 million years ago. This shark was estimated to be 8 to 9 meters (26-30 feet) long. This suggests that the Cardabiodontidae family might have gotten smaller over time. Their teeth also became stronger, and they had fewer small bumps (cusplets) on their teeth.
What Cardabiodon Looked Like

C. ricki was estimated to be up to 5.5 meters (18 feet) long. This was figured out by comparing its backbones to those of a modern great white shark. C. venator was a bit smaller, around 5 meters (16 feet) long, based on its teeth. This makes Cardabiodon one of the largest sharks known from that time.
Shark fossils are rare because their bodies are mostly made of soft cartilage. Cartilage doesn't usually turn into fossils. However, their teeth and sometimes hardened backbones can be preserved.
The skin of Cardabiodon was covered in tiny, teardrop-shaped scales. These scales had grooves and ridges. Such scales help sharks swim faster by reducing drag in the water. This means Cardabiodon was likely a very fast swimmer. It could chase after quick prey.
Cardabiodon's backbones were very strong. They were thicker than those of other large predatory sharks. This suggests that Cardabiodon was a very heavy and stocky shark. Its backbone was stiff and shaped like a spindle. This shape would make it less flexible but very efficient for fast swimming. Cardabiodon also had a larger body compared to the size of its teeth.
Cardabiodon's Teeth

Cardabiodon had different types of teeth in its mouth, which is called being "heterodontic." This means its teeth varied in shape and size depending on where they were in the jaw. Key features of Cardabiodon teeth include strong roots, sturdy crowns, and small bumps called lateral cusplets. The cutting edges of its teeth were smooth, not serrated.
Cardabiodon teeth were moderately sized. The largest teeth from C. ricki were about 3.67 centimeters (1.4 inches) tall. Some lower back teeth were even larger, up to 3.98 centimeters (1.6 inches) tall. C. venator teeth were slightly smaller but thicker. C. venator also had much shorter lateral cusplets than C. ricki.
Scientists have tried to figure out the exact arrangement of Cardabiodon's teeth. They found that Cardabiodon had mainly four types of teeth: symphysial (front middle), anteriors (front), intermediates (between front and side), and lateroposteriors (side and back).
What was special about Cardabiodon's teeth is how their size changed. In its lower jaw, the tooth size suddenly got much bigger at the first lateroposterior tooth. This is different from most mackerel sharks, where teeth usually get smaller towards the back of the mouth. Also, the lower teeth of Cardabiodon were wider than its upper teeth. For most mackerel sharks, the upper teeth are wider.
How Cardabiodon Lived
Growing up

Like modern mackerel sharks, Cardabiodon grew a new ring on its backbones every year. Scientists can count these rings to find out how old the shark was. However, it's hard to find well-preserved Cardabiodon backbones.
Scientists studied the backbones of three young C. ricki sharks and a tooth from a newborn. They found that C. ricki babies were smaller than other ancient sharks at birth. They grew about 29.9 centimeters (11.8 inches) each year. The young sharks studied lived to about 13 years old. But scientists think Cardabiodon probably lived much longer than that.
Around 5 to 7 years of age, the growth rate of C. ricki changed. This suggests that the sharks reached sexual maturity, meaning they could start having their own babies, at this age.
Where Cardabiodon Lived
Cool water shark
Cardabiodon fossils have been found in Canada, the central United States, England, Western Australia, and Kazakhstan. These places were all in cooler, temperate zones, between 40° and 60° latitude, during the Late Cretaceous. This pattern is similar to the modern porbeagle shark, which also prefers cooler waters. This suggests Cardabiodon had an "antitropical distribution," meaning it lived in temperate zones in both the northern and southern hemispheres.
A study in 2010 confirmed this idea. It found that Cardabiodon lived in waters with temperatures between 17.5 and 24.2 degrees Celsius (63.5-75.6°F). The study also noted that the fossils found farthest north were from just before a very warm period about 93 million years ago. This suggests that Cardabiodon might have moved north to find cooler waters as the Earth warmed up.
It's possible that Cardabiodon could travel between hemispheres by swimming through deeper, cooler ocean waters. Modern basking sharks do this. They follow cool paths in deep tropical waters to migrate. The discovery of Cardabiodon teeth in deep-water areas supports this idea.
Ocean home

Cardabiodon likely lived in temperate, offshore waters that were not too deep. Most of its fossils are from North America. In North America, C. venator fossils are often found in the southern part of the Western Interior Seaway. This was a large inland sea that covered much of central North America. This area had a warmer temperate climate.
Even though Cardabiodon preferred cooler waters, it lived during a time when the Earth was warming up. This warming led to new types of animals appearing, like mosasaurs. In the Western Interior Seaway, Cardabiodon lived alongside many other marine animals. As a top predator, it might have hunted them.
These animals included other sharks like Cretodus, Cretalamna, and Cretoxyrhina. There were also large bony fish such as Xiphactinus, seabirds, and marine reptiles. These reptiles included plesiosaurs, sea turtles, and dolichosaurids.
In Western Australia, Cardabiodon shared its habitat with other sharks like Squalicorax and Archaeolamna. Fragmentary remains of pliosaurs were also found there. Other parts of Western Australia were home to ichthyosaurs like Platypterygius.
During this time, there were not many animals living on the ocean floor. This was mainly because of low oxygen levels in the water. This low oxygen event caused many marine invertebrates to die out.
Shark nurseries
Like many modern sharks, Cardabiodon likely used special nursery areas. These were shallow waters where they could give birth and raise their young. These areas offered protection from other predators.
One such nursery area has been found near Mosby, Montana, in the Carlile Shale. Many fossils of young Cardabiodon sharks have been discovered there. Other places in the Western Interior Seaway, like Alberta and Kansas, have also yielded juvenile fossils. Because young Cardabiodon fossils are rare elsewhere, scientists think the entire Western Interior Seaway might have been a large nursery area for this shark.
Images for kids
-
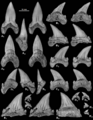
Holotype and paratype teeth of Cardabiodon venator from the Fairport Member of the Carlile Shale in Mosby, Montana.
-
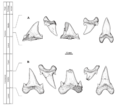
Comparison of the dental characteristics and temporal ranges of the type specimens of C. ricki and C. venator.
-
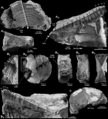
Vertebral centra from the C. ricki holotype, with arrows and dots pointing to growth rings.
-

Reconstructed lower dentition of the C. ricki holotype.
-

Tooth of a juvenile C. ricki from the Haycock Marl in West Australia.
-

Early Cretaceous map showing the localities of Cardabiodon fossils.
See also
 In Spanish: Cardabiodon para niños
In Spanish: Cardabiodon para niños
