
American marten facts for kids
Quick facts for kids American marten |
|
|---|---|
 |
|
| Conservation status | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Carnivora |
| Family: | Mustelidae |
| Genus: | Martes |
| Species: |
M. americana
|
| Binomial name | |
| Martes americana (Turton, 1806)
|
|
| Subspecies | |
|
|
 |
|
| American marten range (note: map is missing distribution in New England and New York State) | |
| Script error: The function "autoWithCaption" does not exist. | |

Script error: No such module "Check for conflicting parameters".
The American pine marten (Martes americana), also called the American marten, is a small mammal from North America. It belongs to the family called Mustelidae, which includes weasels and otters. Sometimes, people just call it the "pine marten." This name comes from a similar animal in Europe. The American marten is smaller and lighter in color than its relative, the fisher.
Contents
What is an American Marten?
The American marten is a long, slender weasel. It's about the size of a mink. It has large, round ears, short legs, and a bushy tail. Its head is somewhat triangular with a sharp nose.
Its fur is long and silky. The color can be light yellowish-brown to dark brown or almost black. The head is usually lighter than the body. Its legs and tail are darker. American martens often have a special patch of fur on their throat and chest. This "bib" can be pale yellow to bright orange.
Male martens are bigger than females. Males are about 15% longer and can be up to 65% heavier. An adult marten is usually 1.5 to 2.2 feet (0.5–0.7 m) long. Its tail is about 5.4 to 6.4 inches (135–160 mm) long. They weigh between 1.1 and 3.1 pounds (0.5–1.4 kg). Martens don't have much body fat. In winter, they might go into a light sleep called torpor to save energy.
Where Do American Martens Live?
American martens live across northern North America. You can find them from Alaska and Canada all the way south to northern New Mexico. They also live from Newfoundland in the east to Humboldt County, California in the west.
In Canada and Alaska, they are found almost everywhere. In the western United States, they mostly live in mountain ranges. Over time, their numbers have changed in some areas. They have disappeared from some places but have also returned to others, like the Great Lakes region. People have even brought them back to areas where they had vanished.
Martens were once thought to live only in old evergreen forests. But studies show they live in many types of forests. This includes old and young forests, both evergreen and leafy (deciduous) trees. They also live in mixed forests. You can find them in the Pacific Northwest, New England, and the Rocky Mountains. They also live in parts of the Midwestern United States, like Wisconsin and Minnesota.
Trapping and losing their forest homes have reduced their numbers. However, they are still more common than the larger fisher. The Newfoundland pine marten (a type of American marten) is considered threatened. The Humboldt marten in the Pacific Northwest is even more endangered. Only a few hundred of them are left.
How Much Space Do They Need?
American martens don't live in very crowded areas. Their population density is low for their size. This means there aren't many martens in one square kilometer. If there isn't much food, their numbers can be even lower.
The size of a marten's home range can vary a lot. It depends on things like their sex, the time of year, and how much food is available. It also depends on the quality of their habitat. Males usually have larger home ranges than females. This might be because females need specific places for dens or food.
Martens mark their home ranges with scent. Male martens often have scars on their heads and shoulders. This suggests they fight with other males over their territory. Males usually don't share their home ranges. But males and females, or adults and young martens, might share space.
Their home range borders often follow natural features. These can be things like forest edges, ridges, lakeshores, or rivers. Open areas like large meadows might also mark their boundaries.
How Do American Martens Behave?
American martens are more active in summer than in winter. In summer, they might be active for up to 60% of the day. In winter, this can drop to as little as 16%. This reduced activity in winter might be because of the cold or less food.
They can be active during the day or night. Their activity patterns can change based on what their prey animals are doing. It also depends on the season and how much food they can find. In some areas, they are active during the day in summer. But in winter, they are mostly active at night.
The distance they travel each day also varies. It depends on their age, sex, and how good their habitat is. It also depends on the weather and how much food is around. Young martens often travel farther each day than adults.
Weather and Snow
Weather can affect how active martens are. It also influences where they rest and how much food is available. They might become inactive during storms or very cold weather. When it's extremely cold, they might rest in places under the snow. This helps them stay warm.
Snow is very important for American martens in many areas. It gives them warmth and places to hunt and rest. They can travel a lot under the snow. Martens are well-suited for snow. Their feet don't sink deep into the snow, even when it's very deep.
Deep snow is especially important where martens live near fishers. Fishers might compete with martens for food or even hunt them. Martens prefer areas with deep snow. Fishers prefer areas with shallow snow. This helps them avoid each other.
Where snow is deep, martens like places where the snow doesn't pack too hard. They also look for structures near the ground that let them get under the snow. Even though they like deep snow, they might hunt more in patches where the snow is a bit shallower. This makes it easier to dig for food.
How Do American Martens Reproduce?
American martens can start having babies when they are about 1 year old. But they might not successfully breed until they are 2 years old. Females can have babies even when they are 15 years old in zoos. In the wild, females as old as 12 have had young.
Adult martens usually live alone. They only come together during the breeding season. Females can have several periods of heat. They usually enter estrus (ready to breed) in July or August. Courtship lasts about 15 days.
The fertilized eggs don't start growing right away. This is called delayed implantation. The actual pregnancy lasts about two months. Females give birth in late March or April. A litter can have 1 to 5 babies, called kits.
Raising the Young
Females use dens to give birth and keep their kits safe. These are called natal dens. Later, they might move their kits to other dens, called maternal dens. Marten mothers use many different places for dens. These include hollow trees, old stumps, logs, piles of wood, or rock piles. They might also use nests or food storage areas of red squirrels.
Females line their natal dens with grass, moss, and leaves. They often move their kits to new dens when the kits are 7–13 weeks old. Mothers spend a lot of time taking care of their kits. As the kits get older, the mother spends less time at the den. Male martens do not help raise the young.
Kits start eating solid food around 42 days old. They come out of the dens when they are about 50 days old. When they first come out, they are active but a bit clumsy. They become more coordinated by 12 to 15 weeks. Young martens reach adult weight around 3 months old.
Kits usually stay with their mother until the end of their first summer. Most of them leave to find their own homes in the fall. The time they leave can vary. Some might leave in early spring. Young martens can travel long distances to find a new home range.
What Do American Martens Eat?
American martens eat whatever food is available. They are opportunistic predators. This means they will eat what they can find easily. They need about 80 calories a day when resting. This is like eating about 3 voles.
Voles are a very important part of their diet. But larger prey, like snowshoe hares, can also be important, especially in winter. They often eat red-backed voles. They also eat meadow voles more than expected. Deer mice and shrews are eaten less often. But they can be important if there isn't other food.
In some coastal areas, fish can be an important food. On the Queen Charlotte Islands, birds were the most important food.
Their diet can change with the seasons or from year to year. In summer, they eat more fruits, plants, and insects. Their diet is usually more varied in the eastern and southern parts of their range. But it's less varied in very cold northern areas.
American martens can also help spread seeds. Seeds often pass through their bodies without being digested. These seeds can then grow into new plants. One study found that blueberry seeds grew better after passing through a marten's gut. Martens can spread seeds over long distances.
How Long Do American Martens Live?
American martens can live up to 15 years in zoos. In the wild, the oldest marten found was 14.5 years old. How long they live depends on where they live, if they are trapped, and the quality of their habitat.
Who Hunts American Martens?
American martens can be hunted by birds of prey and other meat-eating animals. They try to avoid open areas. This might be because they are afraid of predators. The specific predators depend on the area.
In Newfoundland, red foxes are the most common predators. Coyotes and even other martens can also kill them. In British Columbia, most martens are killed by birds of prey. In Oregon, bobcats, birds of prey, and coyotes are common predators. Other animals that hunt martens include:
- great horned owls
- bald eagles
- golden eagles
- Canada lynx
- mountain lions
- fishers
- wolverines
- grizzly bears
- American black bears
- grey wolfs
Most predator attacks happen between May and August. Very few happen in winter.
Trapping and Conservation
The fur of the American marten is very valuable. It looks like the fur of the sable. In the early 1900s, their numbers dropped a lot because of the fur trade. Companies like the Hudson's Bay Company traded their pelts.
Many efforts have been made to protect them and bring them back to areas where they disappeared. This has helped their numbers grow. However, deforestation (cutting down forests) is still a problem for them.
American martens are still trapped for their fur in most places where they live. Trapping can cause many deaths in some marten populations. It can affect how many martens there are, how many males and females, and their ages. Young martens and males are more likely to be trapped. Trapping can be especially harmful in forests where a lot of logging happens.
Other Dangers
Other things that can cause martens to die include drowning, starvation, and getting too cold. They can also choke or get infections from injuries. If they get wet in cold weather, they can die easily.
Martens can also get parasites and diseases. Some diseases have caused many martens to die in certain areas, like Newfoundland.
![]() This article incorporates public domain material from the United States Department of Agriculture document "Martes americana".
This article incorporates public domain material from the United States Department of Agriculture document "Martes americana".
Images for kids
-

In Sitka, Alaska.
-

American martens have a long bushy tail.
-
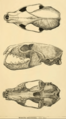
Skull of an American marten.
-

An American marten showing its light-colored throat bib.
-

American martens are active even in winter snow.
-

A young American marten.
.jpg)





See also
 In Spanish: Marta americana para niños
In Spanish: Marta americana para niños
