
Sora (bird) facts for kids
Quick facts for kids Sora |
|
|---|---|
.jpg) |
|
| Conservation status | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Aves |
| Order: | Gruiformes |
| Family: | Rallidae |
| Genus: | Porzana |
| Species: |
P. carolina
|
| Binomial name | |
| Porzana carolina (Linnaeus, 1758)
|
|
 |
|
| Script error: The function "autoWithCaption" does not exist. | |
| Synonyms | |
|
Rallus carolinus Linnaeus, 1758 |
|

Script error: No such module "Check for conflicting parameters".
The sora (Porzana carolina) is a small waterbird that belongs to the Rallidae family. People sometimes call it the sora rail or sora crake. Its scientific name, Porzana, comes from words used in Venice for small rails. The name "Carolina" refers to the Carolina Colony. The word "Sora" likely comes from a Native American language.
Contents
What the Sora Looks Like
Adult soras are about 19 to 30 centimeters (7.5 to 12 inches) long. They have brown feathers on their backs with dark marks. Their face and belly are blue-grey, and their sides have black and white stripes. They have a short, thick yellow beak. There are black marks on their face near the beak and on their throat.
Male and female soras look alike. However, young soras do not have the black face marks. Their faces are whitish, and their chests are buff-colored. Soras usually weigh between 49 and 112 grams (1.7 to 4 ounces). Their wingspan, which is how wide their wings are when spread out, is about 35 to 40 centimeters (14 to 16 inches).
Where Soras Live
Soras live and raise their young in marshes across much of North America. They build their nests in hidden spots among thick plants. The female sora usually lays 10 to 12 eggs, but sometimes as many as 18. The nest is a cup made from marsh plants.
The eggs do not all hatch at the same time. Both parent birds incubate the eggs, meaning they sit on them to keep them warm. Both parents also feed the young birds. Baby soras leave the nest soon after they hatch. They can fly within about a month.
Plants in Sora Habitats
Soras are often found in areas with certain types of plants. These include cattails (Typha species), sedges (Carex species), bulrushes (Scirpus species), smartweeds (Polygonum species), and rushes (Juncus species). They also like rice cutgrass (Leersia oryzoides) and barnyard grasses (Echinochloa species).
Sometimes, soras are seen in farm fields during their migration. For example, a sora was once seen in a cultivated field in Iowa, about 3 kilometers (1.9 miles) from a marsh. Another sora was seen near a large wetland in a soybean field. Soras have also been found on farms in Saskatchewan that mainly grew wheat.
Water Levels and Sora Homes
Soras use areas with different water depths. They are often seen in water less than 30 centimeters (1 foot) deep. However, in Arizona, areas heavily used by soras had an average water depth of 51 centimeters (20 inches). Sora nesting sites often have shallower water than other random spots in the marsh.
If water levels change a lot, soras might leave their nests. For instance, a sora nest with 7 eggs was abandoned in Colorado when the water level rose more than 20 centimeters (8 inches). Soras tend to avoid open water areas. Their numbers are lower in wetlands with more open water.
Vegetation Density and Height
Sora nesting sites usually have more emergent vegetation (plants growing out of the water) than random spots. Wetlands with more live emergent plants tend to have more soras. The density of these plants can vary. Some areas have about 122 stems per square meter, while others have 333 stems per square meter.
The height of the plants in sora habitats also varies. It can range from 20 to 28 centimeters (8 to 11 inches) in some areas to 213 centimeters (84 inches) in others. In western New York, nesting sites had shorter vegetation than other places.
Preferred Plant Types
Soras seem to prefer certain types of plants. In Arizona, they used southern cattail (Typha domingensis) much more than its availability. They also liked bulrushes and a mixed-shrub community. However, they tended to avoid plants like saltcedar (Tamarix chinensis) and arrowweed (Pluchea sericea). They also avoid areas dominated by purple loosestrife.
In Maine, soras used wetlands with more ericaceous vegetation, like leatherleaves (Chamaedaphne species) and sweetgales (Myrica species). In Iowa, broadleaf arrowhead (Sagittaria latifolia) was often found in sora territories. In Wisconsin, soras were more often found in cattail (Typha species) areas than in sedge areas.
Sora Migration
Soras migrate to warmer places for winter. They travel to the southern United States, the Caribbean, and northern South America. The sora is very rarely seen in western Europe. There, it might be confused with the spotted crake. However, the spotted crake always has spots on its chest, a striped crown, and different wing patterns.
Soras look for food by walking or swimming. They are omnivores, meaning they eat both plants and animals. Their diet includes seeds, insects, and snails. Soras are often heard more than seen, but sometimes you can spot them walking near open water. They are quite common, even though their suitable habitat has decreased recently. Their call is a slow, whistled ker-whee, or a descending whinny. Playing sora calls can help you hear or even see them, as they often come to investigate the sound.
Where Soras Are Found
Soras live across most of North America. They breed from Nova Scotia in Canada, northwest to southern Yukon and Northwest Territories. In the US, they breed south to California, Arizona, and New Mexico, and northeast to Pennsylvania and New England.
During winter, soras can be found in the Caribbean and northern parts of South America, including Ecuador, Colombia, and Venezuela. They also winter north through Central America and Mexico to southern California and the coastal southeastern US. In some areas, like central California and parts of Arizona and New Mexico, soras might stay all year.
Spring and Fall Migration
Soras fly north in spring, mostly in April and May. For example, in east-central Kansas, more soras were seen from April 24 to May 7. In southeast Missouri, soras were observed from March 25 to May 6. They were first seen in April or early May in Colorado, Iowa, and Minnesota.
Soras leave their breeding grounds as early as July and as late as October. They start returning to their wintering grounds in Arizona by late July. Most migration happens in August and September. In northern Ohio, the number of soras increased in late August and September due to migrating birds.
Sora Nesting and Life Cycle
Sora nesting activities have been seen from late April through early August. However, most nesting happens from May to early July. In New York, nesting started in late April.
Female soras begin building saucer-shaped nests on the ground or on a platform over shallow water when they start laying eggs. A group of eggs laid at one time is called a clutch. Clutch sizes are usually 8 to 13 eggs, but up to 16 have been reported. Both parents sit on the eggs to keep them warm. This incubation lasts about 19 days.
The eggs hatch over a period of 2 to 13 days. Baby soras are called precocial, which means they are quite developed at birth. They can walk and swim short distances (less than 1 meter or 3 feet) by the end of their first day. Young soras become independent at about 4 weeks old. Soras usually raise one group of young per season.
Sora Survival and Home Range
Not much information is available on how long soras live. Some studies suggest that radio-marked soras might have a lower survival rate, possibly because of the tracking devices. They can be caught by predators or hit by cars.
The size of a sora's home range (the area it uses) can vary. In northwestern Iowa, areas where soras raised their young averaged about 0.2 hectares (0.5 acres). In Arizona, a sora's home range was about 0.6 hectares (1.5 acres) in the early breeding season and over 0.8 hectares (2 acres) after breeding.
Sora Population Density
The number of soras in an area can vary. Some places have up to 12 soras per acre in Colorado, while others have about 0.47 pairs per hectare in Indiana. In Iowa, the average density over two years was 1.3 pairs per hectare.
Soras can be found in marshes of all sizes. However, they might be more common in medium-sized marshes. In Maine, soras used 10% of small wetlands (1 hectare or 2.5 acres), 40% to 50% of medium wetlands (1 to 2 hectares or 2.5 to 5 acres), and 20% of very large wetlands (over 20 hectares or 50 acres). In western New York, soras were more common in marshes that were 40 to 100 hectares (100 to 250 acres) in size.
Soras also seem to like "edge habitats," which are areas where different types of habitats meet. Sora territories were often closer to habitat edges than those of other birds like the Virginia rail.
What Soras Eat

Soras eat many different kinds of food. They commonly eat snails, crustaceans, spiders, and insects. Their insect diet mainly includes beetles, grasshoppers, flies, and dragonflies.
Soras also eat the seeds of many plants. These include smartweeds, bulrushes, sedges, and barnyard grasses. In the eastern United States, they eat seeds from annual wildrice (Zizania aquatica) and rice cutgrass. Other plant foods include crowngrass (Paspalum species) and rice.
Soras eat more plant food in the fall and winter (68%–69% of their diet) than in the spring and summer (40%). For example, plants like hairy crabgrass and fall panicgrass were found much more often in soras' stomachs in the fall than in the spring. In spring, they eat more animal material, mostly adult beetles and snails.
Who Eats Soras?
Sora eggs are eaten by several animals. These include American minks, skunks, coyotes, grackles, crows, and herons. Adult soras can be hunted by American minks, coyotes, and hawks and owls.
Images for kids
-
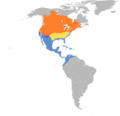
Map showing the distribution of the Sora.
![]() This article incorporates public domain material from the United States Department of Agriculture document "Porzana carolina".
This article incorporates public domain material from the United States Department of Agriculture document "Porzana carolina".
See also
 In Spanish: Polluela sora para niños
In Spanish: Polluela sora para niños
