
Dromaeosaur facts for kids
Quick facts for kids DromaeosaursTemporal range: Upper Jurassic–Upper Cretaceous
|
|
|---|---|
 |
|
| Deinonychus (large) and Buitreraptor Field Museum of Natural History, Chicago |
|
| Scientific classification | |
| Kingdom: | |
| Class: | |
| Superorder: | |
| Order: | |
| Suborder: | |
| Infraorder: | |
| Family: |
†Dromaeosauridae
|
| Subfamilies | |
|
|




Dromaeosaurs are a group of dinosaurs that were a lot like birds. They were theropod dinosaurs, which means they walked on two legs. This group includes the famous Velociraptor.
Dromaeosaurs were small to medium-sized meat-eaters (carnivores). They had feathers and lived during the Cretaceous period. People often call them raptors. This name became very popular after the movie Jurassic Park. They were very fast hunters.
These dinosaurs lived all over the world. The first dromaeosaurs appeared about 167 million years ago (mya) in the Middle Jurassic period. Larger types showed up later, around 124 mya, in the Lower Cretaceous period. These bigger ones have only been found in the northern part of the world. Dromaeosaurs lived until the end of the Cretaceous period, about 65.5 mya. This was when a big event, like an asteroid impact, caused many species to go extinct.
Scientists know dromaeosaurs existed in the Middle Jurassic because they have found fossil teeth from that time. However, full body fossils from that early period are rare.
Contents
What Did Dromaeosaurs Look Like?
The bones of dromaeosaurs tell us they were active and fast runners. They were also very closely related to birds.
Dromaeosaurs had a fairly large head. Their teeth were serrated, like a saw, and their snout was narrow. Their eyes faced forward, which helped them see in 3D (called binocular vision). This is useful for hunting.
Like most other theropods, dromaeosaurs had an S-shaped neck. Their body was short but deep. They had long arms and big hands with three long fingers. Each finger ended in a large, sharp claw.
Their hip bones had a special part called a 'pubic boot'. This part stuck out under their tail. It was where strong muscles and tendons attached, helping them move.
Dromaeosaurs had a large, curved claw on their second toe. Their tails were thin and mostly used for balance when they ran.
Many, and probably all, dromaeosaurs were covered in feathers. This included big feathers on their wings and tails, and softer, down-like feathers on their bodies.
Their Special Foot
Like other theropods, dromaeosaurs walked on their two back legs. But unlike most other theropods, dromaeosaurs usually held their second toe off the ground. This means they walked on only their third and fourth toes.
The second toe had a very large, curved, sickle-shaped claw. Scientists think this claw was used to catch prey. Smaller dromaeosaurs might have also used it for climbing trees.
One type of dromaeosaur, Balaur bondoc, had two special claws on each foot. Both its first and second toes were held up and had big, curved claws.
Their Long Tail
Dromaeosaurs had long tails. Many of their tail bones had bony, rod-like parts. Some also had bony tendons. These features made the tail stiff, so it mostly bent only at its base. This meant the whole tail could move like a single, strong lever.
However, one well-preserved Velociraptor fossil shows a tail that could bend sideways in an S-shape. This suggests their tails were somewhat flexible. It's thought the tail helped them balance and steer while running. For Microraptor, a fan of feathers at the end of its tail might have helped it glide and steer in the air.
How Big Were They?

Dromaeosaurs came in different sizes. The smallest, like Mahakala, were about 0.7 meters (2.3 feet) long. The largest, like Utahraptor and Achillobator, could be over 6 meters (20 feet) long. Some Utahraptor fossils suggest they might have been even bigger, up to 11 meters (36 feet) long!
It seems that large size evolved more than once in dromaeosaurs. Scientists have also found teeth from a giant dromaeosaur on the Isle of Wight in England. This dinosaur was as big as Utahraptor but had teeth similar to Velociraptor.
Mahakala is the smallest and most basic dromaeosaur found so far. This, along with other small feathered dinosaurs like Microraptor and Anchiornis, suggests that the common ancestor of dromaeosaurs, troodontids, and birds might have been very small, perhaps around 65 cm (2 feet) long and weighing less than a kilogram.
Their Feathers

Fossils clearly show that dromaeosaurs were covered in feathers. Some fossils even show long feathers on their hands and arms (like wing feathers) and tails, plus shorter, soft feathers on their bodies.
Other fossils don't show actual feather marks, but they have bumps on the arm bones. These bumps are where strong wing feathers would have attached, just like in some birds today. This feather pattern looks a lot like Archaeopteryx, an early bird.
The first dromaeosaur found with clear evidence of feathers was Sinornithosaurus in China in 1999.
Many other dromaeosaur fossils have been found with feathers, some even with fully formed feathered wings. Microraptor even had a second pair of wings on its back legs! Even if direct feather marks aren't seen, some fossils have "quill knobs." These are small bumps on the bones where feathers attached. Rahonavis and Velociraptor fossils have these knobs, proving they had feathers too.
Because of this, scientists believe that even the larger dromaeosaurs that lived on the ground had feathers. Even birds today that can't fly still have most of their feathers.
How Dromaeosaurs Lived
The Sickle Claw's Use

Scientists have debated what the big "sickle claw" on the second toe was for. When John Ostrom first described it for Deinonychus in 1969, he thought it was a slashing weapon. He believed dromaeosaurs used powerful kicks to cut into their prey, similar to how some saber-toothed cats used their teeth. This idea was applied to all dromaeosaurs.
Another idea is that the claw helped them climb onto bigger prey to hold it down, and also acted as a stabbing weapon.
Ostrom compared Deinonychus to modern birds like the ostrich and cassowary. Cassowaries have very long claws (up to 125 mm or 5 inches) and can cause serious injury with them. The seriema bird also has an enlarged second toe claw, which it uses to tear apart small prey.
Scientists have also studied the curve of the dromaeosaur "sickle claw" and compared it to claws of modern birds and mammals. They found that the amount of curve in a claw often shows how an animal lives. Very curved claws usually belong to climbing animals, while straighter claws are for animals that live on the ground. The sickle-claws of Deinonychus were very curved (160 degrees), which is typical for climbing animals. Their front limb claws were also curved for climbing.
Paleontologist Peter Mackovicky noted that small, early dromaeosaurs like Microraptor probably climbed trees. But he wondered why huge dromaeosaurs like Achillobator still had curved claws if they were too big to climb trees.
Living in Groups

Fossils of Deinonychus have been found in small groups near the bones of a large plant-eating dinosaur called Tenontosaurus. Some scientists thought this meant dromaeosaurs hunted in groups, like some modern mammals. However, not all scientists agreed. A study in 2007 suggested that Deinonychus might have just gathered around a kill in a disorganized way, rather than hunting together.
The first large set of dromaeosaur footprints was found in China. These tracks were made by a large dromaeosaur, similar in size to Achillobator. The tracks showed that the sickle-claw was held off the ground. Six dromaeosaurs of similar size walked together along a shoreline. They were spaced about one meter apart and moved in the same direction at a slow pace. These tracks show that some dromaeosaur species lived in groups. While these specific tracks don't show hunting, it's still possible that groups of dromaeosaurs hunted together.
Flying and Gliding

Scientists have suggested that at least two types of dromaeosaurs could fly or glide. One, Rahonavis, was first thought to be a bird. Later studies showed it was a dromaeosaur. It might have been able to fly actively. The front limbs of Rahonavis were very strong, even stronger than Archaeopteryx, and had strong attachments for muscles needed for flapping flight. Scientists believe Rahonavis could probably fly, but it might have been clumsy compared to modern birds.
Another dromaeosaur, Microraptor, might have been able to glide. It had well-developed wings on both its front and back legs. A 2005 study suggested that Microraptor's wings worked like a "biplane" (an airplane with two sets of wings). It likely glided by launching from a high spot, swooping down in a U-shape, and then lifting up to land on another tree. Its tail and back wings would have helped it control its position and speed. The study also found that Microraptor had the basic features needed for sustained flight, not just gliding.
Dromaeosaurs and Birds
Their Link to Birds

Dromaeosaurs share many features with early birds. Scientists are still discussing exactly how they are related.
Some scientists believe dromaeosaurs are very closely related to birds. Others think birds are more like cousins to dromaeosaurs and another group called troodontids.
Today, most paleontologists agree that dromaeosaurs are most closely related to troodontids. Together, these two groups form the Deinonychosauria. This larger group is the closest relative to birds.
Scientists generally agree that there isn't enough evidence yet to say for sure if any dromaeosaurs could fly or glide, or if they evolved from ancestors that could.
Other Ideas About Flight

Some researchers have suggested that dromaeosaurs might have come from flying ancestors.
In 2002, a study found that Microraptor was a very early type of dromaeosaur. Because Microraptor had feathers and wings, some scientists thought that the first dromaeosaur might have been able to glide. If this is true, then the larger dromaeosaurs would have later lost the ability to glide as they evolved to live on the ground.
A few researchers, like Larry Martin, believe that dromaeosaurs and all similar dinosaurs are not true dinosaurs at all. Martin used to say that birds were not related to these dinosaurs. But in 2004, he changed his mind and agreed they are very close relatives. Martin now thinks that these dinosaurs were actually birds that lost the ability to fly. He believes birds evolved from non-dinosaur ancestors.
However, a challenge to these ideas came in 2007 when a new dromaeosaur, Mahakala, was discovered. Scientists found that Mahakala was the most basic and earliest member of the Dromaeosauridae family, even older than Microraptor.
Mahakala had short arms and could not glide. This suggests that the first dromaeosaur could not glide or fly. Based on this, scientists think that not being able to fly was the original state for dromaeosaurs.
Dromaeosaur Family Tree

The family Dromaeosauridae was named in 1922, with only one new dinosaur called Dromaeosaurus. Dromaeosauridae and Troodontidae together form the group Deinonychosauria.
The Dromaeosauridae family was officially defined in 1998 as a group that includes Dromaeosaurus but not Troodon, Ornithomimus, or Passer (a type of bird). The different smaller groups within Dromaeosauridae, called "subfamilies," have also been defined.
The exact organization of these subfamilies is still being worked out. For example, Mahakala, which is a very basic dromaeosaur, doesn't fit neatly into any of the existing sub-groups. The most basic subfamily of dromaeosaurs is often thought to be the Unenlagiinae.
All known dromaeosaur skin impressions come from this group. They all show a lot of feathers and well-developed wings. Some species in this group might have even been able to fly actively.



Here is a list of some of the dromaeosaur groups and their members:
- Family Dromaeosauridae
- Subfamily Dromaeosaurinae
- Subfamily Microraptorinae
- Cryptovolans
- Graciliraptor
- Hesperonychus
- Microraptor
- Subfamily Saurornitholestinae
- Atrociraptor
- Bambiraptor
- Saurornitholestes
- Subfamily Unenlagiinae
- Subfamily Velociraptorinae
Velociraptorinae is a group within the Dromaeosauridae family. They lived from the Late Jurassic to the end of the Cretaceous period. This group includes Velociraptor, Deinonychus, Tsaagan, Saurornitholestes, and Balaur. Scientists have also found teeth from a giant velociraptorine, as big as Utahraptor, on the Isle of Wight in England. Generally, velociraptorines have longer, narrower jaws and more slender bodies than dromaeosaurines.
-
-
- Adasaurus
- Balaur
- Deinonychus
- Linheraptor
- Nuthetes
- Tsaagan
- Velociraptor
-
Dromaeosaurs in Movies and Books
A Deinonychus appeared in John Brosnan's 1984 book Carnosaur and its movie version.
Velociraptor, a dromaeosaur, became very famous after the 1993 Steven Spielberg movie Jurassic Park. However, the Velociraptor in the movie was much larger than the real Velociraptor dinosaurs. Robert Bakker, a paleontologist, said that Spielberg was disappointed with the size of Velociraptor and made it bigger for the movie. Soon after, Bakker named Utahraptor, which was closer to the size shown in the film, or even larger.
Michael Crichton used the name Velociraptor for the much larger raptor in his books, which the first two movies were based on. The way dromaeosaurs were shown in the original Jurassic Park movie was accurate for its time, but now we know some things were wrong, like the lack of feathers. Jurassic Park III tried to fix this by adding quill-like structures around the heads of some dromaeosaurs. However, these didn't look like the actual feathers found on dromaeosaur fossils.
Dromaeosaurids also appear in many of The Land Before Time movies, starting with the third one.
Images for kids
-

Life restoration of Deinonychus antirrhopus
-

Size comparison of several dromaeosaurids
-

Diagram comparing three of the largest described genera: Achillobator, Dakotaraptor and Utahraptor
-

Preserved feather traces in a fossil Zhenyuanlong suni.
-

The Thermopolis specimen of Archaeopteryx, which showed that it also had a hyperextendible second toe
-

Luanchuanraptor, a dromaeosaurid of uncertain placement
-

Halszkaraptor, a halszkaraptorine
-

Austroraptor, an unenlagiine
-

Changyuraptor, a microraptorian
-

Saurornitholestes, a eudromaeosaur
-

Life restoration of Deinonychus preying on a Zephyrosaurus using the sickle claw for prey restraint
-
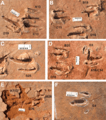
Tracks of the ichnogenus Paravipus didactyloides, interpreted as representing two individuals that were moving in the same direction
-

Microraptor gui fossil with impressions of feathered wings







.jpg)

See also
 In Spanish: Dromeosáuridos para niños
In Spanish: Dromeosáuridos para niños
