
Domestication of the dog facts for kids

The domestication of the dog is the story of how dogs became our companions. It tells us how dogs changed from wolves and became the first pets. Scientists believe that all dogs, old and new, came from an ancient type of wolf that is now extinct. This old wolf was different from the wolves we see today. Dogs look a lot like modern grey wolves because dogs and wolves have mixed over time. The grey wolf is the dog's closest living relative.
Dogs are a type of animal related to wolves. The ancestors of dogs and modern wolves separated about 30,000 to 40,000 years ago. This happened just before or during the time of the Last Glacial Maximum, which was about 20,000 to 27,000 years ago. This separation date is the earliest possible time when dogs started to become domesticated.
One of the biggest changes in human history was when we started to tame animals. This began with wolves and hunter–gatherers over 30,000 years ago. Dogs were the very first animal, and the only large carnivore (meat-eater), that humans ever tamed. Dogs became domesticated because some wolves were less aggressive and more friendly towards humans. These friendly wolves got benefits from being near humans, which helped them survive and have more pups. This process is called social selection.
The oldest clear evidence of a dog is the Bonn-Oberkassel dog, found buried with humans 14,200 years ago. Some older remains, from 36,000 years ago, might also be dogs, but scientists are still debating this.
Dogs were tamed long before humans started farming. Farming began about 11,000 years ago in the Near East. Scientists still argue about where dogs were first tamed. But most evidence points to Eurasia, possibly Central Asia, East Asia, or Western Europe. By the end of the last Ice Age, about 11,700 years ago, different types of ancient dogs had spread around the world. These include dogs found in the Levant (7,000 years ago), Karelia (10,900 years ago), Lake Baikal (7,000 years ago), ancient America (4,000 years ago), and the New Guinea singing dog (still alive today).
In 2021, a review of studies suggested that dog domestication started in Siberia about 19,700 to 26,000 years ago. This happened with ancient people called Ancient North Eurasians. From Siberia, dogs then spread east into the Americas and west across Eurasia. We haven't found very old dog remains from this time and place yet. This is because not many archaeological digs have happened in those areas.
The oldest known dog skeletons were found in the Altai Mountains of Siberia and a cave in Belgium. They are about 33,000 years old. This might mean that dogs were tamed in different places at the same time.
Contents
- How Dogs Became Different from Wolves
- Dog Domestication
- First Dogs
- Images for kids
How Dogs Became Different from Wolves

Genetic studies show that the grey wolf is the closest living relative of the dog. It's hard to figure out the exact family tree of dogs using DNA from modern dogs and wolves. This is because:
- An extinct wolf from the Late Pleistocene era is the true ancestor of dogs, not modern wolves.
- The split between dog ancestors and modern wolves happened very quickly. This makes it hard to pinpoint the exact date.
- Dogs and wolves have bred with each other since dogs were tamed. This mixing of genes makes things more complicated.
- Not enough changes (mutations) have happened in dog DNA since domestication. This also makes dating difficult.
Ancient Wolves
The Late Pleistocene was a time of ice ages and big climate changes. Humans were also moving into new areas. A huge grassy plain called the mammoth steppe stretched across Eurasia and into Alaska. At the end of this period, the climate changed quickly and severely. This led to many large animals, like mammoths, dying out. Wolves, however, survived.
The grey wolf is one of the few large meat-eaters that survived the extinctions of the Late Pleistocene. But their numbers did drop globally. Modern wolves today came from a single group that spread out from a safe place in Beringia. They replaced older wolf groups across Eurasia and North America. This group of wolves probably didn't create dogs. But they did mix with dogs, which gave them new fur colors and helped their immune systems.

The first dogs came from wolf groups that lived during the Late Pleistocene. Fossils show that wolves changed in size and shape during this time. Some ancient wolves had stronger jaws and teeth than modern wolves. This might have helped them eat large prey and bones from the Pleistocene megafauna. These ancient wolves had different DNA than modern wolves, suggesting they died out.
Scientists used to think dogs came from the modern grey wolf. But a 2014 study showed this was wrong. Dogs came from an extinct type of wolf.
It was such a long standing view that the gray wolf that we know today was around for hundreds of thousands of years and that dogs derived from them. We're very surprised that they're not.
When Dogs and Wolves Separated
The date when dogs and wolves separated doesn't mean that's when domestication started. It just gives us the earliest possible time. For example, horses separated from their wild relatives about 45,000 years ago. But they were only tamed about 5,500 years ago.
Recent studies show that dog ancestors and modern wolves separated between 20,000 and 40,000 years ago. This is the earliest possible time for domestication.
In 2015, scientists studied the DNA of an ancient wolf from the Taymyr Peninsula. This wolf lived 35,000 years ago. By using this wolf's DNA, they could figure out that dogs and modern wolves separated between 27,000 and 40,000 years ago. This happened before the coldest part of the last Ice Age. In 2017, another study looked at the DNA from the cell nucleus of three ancient dogs. It found that dogs and wolves separated between 36,900 and 41,500 years ago.
Before this separation, the wolf group that became dogs was larger than other wolf groups. After the split, the dog population became much smaller.
A 2020 study of Eurasian wolves found that they and dogs share a common ancestor from 36,000 years ago. This supports the idea that all modern wolves came from one group that spread after the Last Ice Age. It also supports finding dog-like fossils older than 30,000 years.
Where Dogs and Wolves Separated
Based on Modern DNA
Some genetic studies of modern dogs suggested that dogs came from Southeast Asia or South China. This is because dogs there show more genetic variety. However, this idea has been questioned. Dog fossils have been found in Europe that are 15,000 years old, but only 12,000 years old in far eastern Russia.
Other studies suggested origins in Central Asia or the Middle East. But these studies often compared purebred dogs (which have less genetic variety) with village dogs, which isn't always a fair comparison. Also, some findings were due to dogs and wolves mixing later on. A 2014 study found that no modern wolf group is closer to dogs than any other. This suggests the dog's ancestor is extinct.
Based on Ancient DNA
Most older genetic studies used modern dog breeds and living wolf groups. They made assumptions that might not be true. For example, they assumed modern wolves were the direct ancestors of dogs. More recently, scientists have been able to study ancient DNA from fossils. This new field is called Paleogenomics.
Arctic Siberia

In 2015, a study looked at ancient dog-like animal remains from Zhokhov Island and the Yana River in arctic Siberia. Some of these remains were 28,000 years old and matched the DNA of modern dogs. One 47,000-year-old animal from Duvanny Yar was very similar to modern dogs. The scientists think that ancient Siberian wolves helped create the modern dog's genetic makeup.
Southern Siberia

In 2013, a study examined a well-preserved skull and jaw of a dog-like animal from Razboinichya Cave in the Altai Mountains of southern Siberia. It was dated to 33,300 years ago, making it older than other evidence from Europe and the Near East. Its DNA was more like a dog's than a wolf's. Later, scientists said it was somewhere between a dog and a wolf. In 2017, experts agreed it was an extinct type of dog from an extinct group of small wolves.
Europe

In 2013, a study looked at the DNA of 18 ancient dog-like fossils from Europe and America. It compared them to modern wolves and dogs. The study found that 78% of modern dogs had a genetic link to ancient dog-like animals found in Europe. This suggests dogs might have originated in Europe between 18,800 and 32,100 years ago. This supports the idea that dogs were tamed before farming, near the end of the Last Ice Age, when hunter-gatherers hunted large animals.
The study also found three ancient dog-like animals from Belgium (one from 36,000 years ago, and two from 30,000 and 26,000 years ago). Their skulls had some dog-like features. This might mean that domestication attempts happened more than once, but some failed.
One idea is that dog domestication happened during very cold periods called Heinrich events. These events caused humans to change how they lived. Taming large wolves might have been a way to adapt to the harsh environment.
In 2019, dog remains from Italy, dated 20,000 to 24,700 years ago, were found to be genetically similar to modern dogs. In 2020, even older dog remains (14,000 years old) were found in southern Italy. These were related to the Bonn-Oberkassel dog from Germany. This suggests a common ancestor for these early European dogs around 28,500 years ago.
How Dogs Changed Physically
The first dogs probably looked a lot like wolves. We don't know exactly what physical changes happened when dogs and wolves separated. It's hard to tell the earliest dogs from wolves using their bones. This is because the features that make dogs different (like tooth size or skull shape) weren't fully developed yet.
Fossils suggest that there were both wolf-like dogs and dog-like wolves. If early dogs scavenged human leftovers, then being wolf-like might have been helpful. As humans settled down, dogs might have become smaller and more distinct.
The most widely accepted oldest dog remains are from the Bonn-Oberkassel dog, about 15,000 years old. Older remains (30,000 years old) are still debated as to whether they are dogs or wolves.
Two Ancestries for Dogs in West Asia, Africa, and Southern Europe
New research on 72 ancient wolf genomes (DNA) from the past 100,000 years shows something interesting. Early and modern dogs are more genetically similar to ancient wolves from Asia than from Europe. This suggests that dogs were first tamed in the East.
The research also found that dogs might have two different ancestries. This means two separate groups of wolves gave DNA to dogs.
- Early dogs from northeastern Europe, Siberia, and the Americas seem to have one origin from the eastern source.
- But early dogs from the Middle East, Africa, and southern Europe seem to have some DNA from another source, related to Middle Eastern wolves. They also have DNA from the eastern source.
This could mean that wolves were tamed more than once, and then these different dog groups mixed. Or, it could mean that dogs were tamed once, and then early dogs mixed with wild wolves.
The study also showed how wolf DNA changed over 100,000 years. One gene variant, called IFT88, became very common in wolves over 10,000 years. It's still in all wolves and dogs today. This gene affects how skull and jaw bones develop. It might have spread because wolves with a certain head shape had an advantage when hunting during the Ice Age.
Dog Domestication
Animal domestication is a process where a group of animals changes over time as they adapt to living with another species. Dogs were the first animals tamed by humans. They were domesticated and spread across Eurasia before the end of the Ice Age. This was long before humans started farming or taming other animals. Dogs might have had a big impact on early human history and the growth of civilization.
Scientists have been trying to figure out when and where dogs were first tamed for decades. Genetic studies suggest it started over 25,000 years ago, in one or more wolf groups in Europe, the Arctic, or East Asia. It's clear that dogs came from grey wolves. The wolf groups involved are probably extinct now. Even with many studies, there's still no clear answer about the exact time, place, or number of wolf groups involved.
Around 10,000 years ago, farming began, and people started living in one place. This led to dogs changing physically from their wolf ancestors, including differences in size. Dogs have gone through two major population drops. One happened when they were first tamed, and another when dog breeds were created.
Socialization
Humans and wolves both live in complex social groups. How they first got together is still a mystery. Some scientists think that the word "domestication" might not be right for dogs. This word usually means humans breeding wild animals for food or materials. But dogs are large meat-eaters.
Instead, some think dogs were simply "socialized" to live with humans. Even today, some dogs live with families but are not socialized. They might act aggressively towards strangers, just like a wild wolf. There are also stories of wild wolves approaching people in remote areas, trying to play and form friendships. This suggests that wolves had to become socialized before they could be truly domesticated.
Wolves on Ellesmere Island today don't fear humans much. This is because they rarely see people. They will approach humans carefully and curiously.
Commensal Pathway
Dogs are a classic example of an animal that likely became domesticated by living near humans and sharing their food. This is called a commensal pathway. Dogs were the first animals tamed, and they were widespread before the end of the Ice Age. This was long before farming or other animal domestication. It might have been easier to tame a meat-eater first, as they are less afraid to approach other species. Among meat-eaters, the best choice would be one that doesn't need an all-meat diet, can hunt for itself, is a manageable size, and has a good temperament. Wolves are very social and cooperative animals.
Human Campfire Theory
Ancient DNA supports the idea that dog domestication happened before farming. It started near the end of the Last Ice Age, when hunter-gatherers hunted large animals. Early dogs might have eaten leftovers from human kills. They might have also helped humans hunt or protected them from other predators at kill sites.
Wolves were probably drawn to human campfires by the smell of cooking meat and discarded food. They might have first stayed loosely around the camps. Then, they might have started to see the camps as part of their territory. Their growls would warn humans about outsiders. The wolves most likely to be drawn to human camps were the less aggressive ones. These wolves were less scared, handled stress better, and were less wary of humans. This was the start of a process called self-domestication.
Migratory Wolves Theory

On the mammoth steppe, wolves hunted in packs and shared risks. This put them at the top of the food chain. Some wolves followed large herds of reindeer, making the herd stronger by hunting the weak and sick. These wolves were like the first herders, long before humans. Wolves could keep up with the herds and eat their prey quickly before other predators arrived. One study suggests that during the Last Ice Age, some humans teamed up with these "pastoralist" wolves and learned their hunting methods.
Many early humans were gatherers or specialized hunters. But some adopted the wolves' lifestyle, following herds of reindeer and other hoofed animals. Humans would take the best animals, while the wolves kept the herd healthy. This group of humans became the first herders, and these wolves became the first dogs.
Large carcasses left by human hunter-gatherers might have led some wolves to follow humans as they moved. This could have made them different from wolves that stayed in one area. A closer relationship might have then developed, like hunting together or defending each other from other animals or humans.
Food Sharing Theory
Dogs were the only animals tamed by mobile hunter-gatherers. Humans and wolves were both strong pack hunters of large prey. They competed for food and could even kill each other. One study suggests how humans might have tamed such a dangerous competitor.
Humans are different from other large meat-eaters. Our bodies can only get about 20% of their energy from protein. Eating too much protein can make humans sick. During the harsh winters of the Last Ice Age, plants were not available. Fat and grease were more important than meat for humans. Game meat would have been lean, but bones contained fatty oils.
Wolves, however, can live on a protein-based diet for months. Calculations show that if humans took the fat and oils from animal carcasses, there would still be enough leftover calories to feed proto-dogs or wolves. This means there was no need for competition. Hunting together and protecting each other from other predators would have helped both species, leading to domestication.
Genetic Changes


Coat Color Study
Dogs have many different coat colors and patterns. In many animals, different colors come from a gene called Agouti. This gene controls whether hair follicles make black/brown pigments or yellow/white pigments. The most common wolf coat pattern is agouti, with banded hairs on top and lighter shading underneath. Yellow is a dominant color in dogs and is found in many dogs around the world, including the dingo.
A 2021 study looked at the DNA of dogs and wolves, focusing on coat color. It found that most dog colors were similar to wolf colors. But the dominant yellow color in dogs was closely related to white in arctic wolves from North America. This suggests that dominant yellow in dogs and white in wolves came from a common ancestor. The study suggests that about 35,000 years ago, an extinct type of canid (dog-like animal) mixed its genes with Late Pleistocene grey wolves. This mixing brought new color diversity. This color diversity was seen in wolves 35,000 years ago and in dogs 9,500 years ago.
Dietary Adaptation
Dogs' bodies seem to have changed to handle different diets. First, they adapted to more fat. Later, they adapted to more starch, as they lived closer to humans.
Compared to wolves, the dog's DNA shows changes related to brain function, behavior, and fat metabolism. This ability to process fats was important when early dogs hunted and ate alongside hunter-gatherers. As proto-dogs helped humans hunt, they might have eaten more fat. Also, humans would take the best parts of the carcass, leaving different types of tissues for the dogs. This unique food situation might have led to changes in the dogs' diet.
Behavior
The most important changes in domestication seem to be in social behavior. These changes are linked to genes related to brain function and hormones. Differences in behavior between dogs and wolves might be due to changes in genes linked to human Williams-Beuren syndrome. This syndrome causes increased friendliness, which might have been important during domestication.
A 2014 study of dog and wolf DNA found that dogs' tameness wasn't just less fear. It also showed better synaptic plasticity, which helps with learning and memory. The study suggests that dogs' improved learning and memory helped them be less afraid of humans.
Unlike other tamed animals chosen for things like meat or wool, dogs were first chosen for their behavior. A 2016 study found only 11 genes that were consistently different between wolves and dogs. These gene changes likely came from humans choosing dogs for certain traits. There was evidence of selection for genes that affect the body's stress response, like adrenaline. These genes are involved in how the brain processes emotions and the "fight-or-flight" response. This suggests that the tameness and emotional processing we see in dogs might be due to changes in this pathway. Dogs are generally less fearful and aggressive than wolves. Some of these genes are linked to aggression in certain dog breeds, showing their importance in both early domestication and later breed creation.
Role of Epigenetics
Changes in hormones linked to domestication might be related to epigenetic changes. A study comparing dogs and wolves found 68 differences in how their DNA was marked. These marks are linked to genes that affect how the brain works. For example, a dog's social behavior is linked to a gene called OXTR, which is important for the hormone oxytocin. This change is due to epigenetic marking of the OXTR gene. Differences in DNA marking have been found between wolves and dogs, and between different dog breeds. This means epigenetics might have been important for both dog domestication and the creation of dog breeds.
Like humans, wolves have strong social and emotional bonds within their groups. This might have been the basis for the strong bond between dogs and humans. A 2019 review suggested a new idea called Active Social Domestication. It says that the social environment of dog ancestors caused changes in their brains, leading to quick domestication.
Dog and Human Coevolution
Parallel Evolution
Being the first animal tamed by humans created a strong bond between dogs and humans. Their histories became linked. Dogs went with humans when they moved to new places. They show similar adaptations, like being able to live in high altitudes with low oxygen.
Many genes show similar changes in both dogs and humans. This has led to studying how their genes changed together. For example, genes that affect the brain's serotonin system suggest that coevolution led to less aggressive behavior when living in crowded places.
Dogs also get many of the same diseases as humans, like cancer, diabetes, and heart disease. The way these diseases work in dogs is similar to humans, and they respond to treatment in similar ways.
Behavioral Evidence
Convergent evolution is when different species develop similar solutions to the same problem. For example, fish, penguins, and dolphins all have flippers to move through water. What's special about dogs and humans is that they show psychological convergence. Dogs have developed ways of thinking that are more like humans than even our closest ape relatives. Dogs have special skills for understanding human social cues and communication. These skills seem more flexible and human-like than those of chimpanzees or other great apes. This suggests that both dogs and humans might have developed similar social and communication skills to interact with each other.
Studies show that dogs can follow human pointing gestures. They can also tell how humans are feeling from their facial expressions. Most people can tell from a dog's bark if it's alone, being approached by a stranger, playing, or being aggressive. They can also tell how big a dog is from its growl.
In 2015, a study found that when dogs and their owners make eye contact, levels of the hormone oxytocin increase in both. Oxytocin is known for its role in bonding. This suggests that eye contact helped strengthen the bond between humans and dogs over time.

The dog could have arisen only from animals predisposed to human society by lack of fear, attentiveness, curiosity, necessity, and recognition of advantage gained through collaboration....the humans and wolves involved in the conversion were sentient, observant beings constantly making decisions about how they lived and what they did, based on the perceived ability to obtain at a given time and place what they needed to survive and thrive. They were social animals willing, even eager, to join forces with another animal to merge their sense of group with the others' sense and create an expanded super-group that was beneficial to both in multiple ways. They were individual animals and people involved, from our perspective, in a biological and cultural process that involved linking not only their lives but the evolutionary fate of their heirs in ways, we must assume, they could never have imagined. Powerful emotions were in play that many observers today refer to as love – boundless, unquestioning love.
Human Adoption of Some Wolf Behaviors
... Isn't it strange that, our being such an intelligent primate, we didn't domesticate chimpanzees as companions instead? Why did we choose wolves even though they are strong enough to maim or kill us? ...
In 2002, a study suggested that early humans and wolves might have tamed each other. This partnership would have changed both into humans and dogs. It would have deeply affected human psychology, hunting, and social behavior.
Early humans changed from scavenging to hunting large animals. They started living in bigger, more complex groups. They learned to hunt in packs and cooperate. Since these are also traits of wolves and dogs, it's possible these behaviors became stronger when wolves and humans lived together. Hunting together led to defending each other. Wolves defend their territory, and perhaps humans' sense of territory grew stronger by living with wolves. Forming partnerships has been key to human survival. Strong bonds exist between same-sex wolves, dogs, and humans. Today, the strongest bond between different species is between humans and dogs. The idea of friendship is ancient, but it might have been strengthened by this inter-species relationship, helping both survive.
A 2003 study compared the behavior of chimpanzees, wolves, and humans. Cooperation among chimpanzees is limited. But wolves are very social and cooperative. They hunt together, carry heavy items, and help feed each other's young. These behaviors are similar to human societies. This suggests that social cooperation in canids (dog family) is an older trait than in humans. Today's wolves might even be less social than their ancestors. They no longer have access to huge herds of animals. They live more like coyotes or foxes. Sharing within families might be a trait early humans learned from wolves. Wolves dug dens long before humans built huts. So, it's not clear who tamed whom.
First Dogs
Dogs Tamed in East Asia
A study by Savolainen using mitochondrial DNA suggests that dog domestication began in China or Southeast Asia 33,000 years ago. Then, 18,000 years later, dogs spread from Southeast Asia to Africa and the Middle East. They arrived in Europe about 10,000 years ago, leading to modern dog breeds.
Savolainen noted that many studies that disagreed with an East Asian origin didn't include wolf or dog samples from China or Southeast Asia.
Dogs Tamed in Siberia 23,000 Years Ago
It's hard to find where dogs first came from because we don't have much data on extinct Ice Age wolves. Also, the physical changes between wild and tame populations were small at first. And there isn't much human culture left from that time.
In 2016, a genetic study found that ancient and modern dogs belong to an Eastern Eurasian group and a Western Eurasian group. In 2017, another study found that dogs and wolves separated between 36,900 and 41,500 years ago. Then, Eastern and Western Eurasian dogs separated between 17,500 and 23,900 years ago. This suggests that dogs were tamed only once, between 20,000 and 40,000 years ago.
In 2021, a review of evidence suggested that dogs were tamed in Siberia about 23,000 years ago by ancient North Siberians. Dogs then spread from Siberia with people migrating east into the Americas and west across Eurasia. The ancient North Siberians were people whose remains have been found at sites like Yana RHS (31,600 years ago) and Mal'ta (24,000 years ago) in Siberia. We still need to find ancient dog remains from this time and place to fully support this idea.
The review suggests that the harsh climate of the Last Ice Age might have brought humans and wolves closer. Both species hunted the same prey. Their increased interactions might have led to sharing food, wolves being drawn to human camps, and eventually domestication.
DNA evidence shows that most modern dogs belong to one of four main groups. The largest group, Haplogroup A, first split about 22,800 years ago. This is the oldest known split between any two dog DNA groups. As humans moved across Siberia and into the Americas, their DNA also split. Scientists found a link between human and dog migrations and population splits. This suggests that dogs went wherever people went. By tracing these human and dog lineages, scientists think dogs were first tamed in Siberia almost 23,000 years ago by North Siberians.
Another study looked at the complete DNA of 555 modern and ancient dogs. It showed a population increase around 23,500 years ago, which matches the proposed split of dog ancestors from modern wolves. A much larger population increase happened after 15,000 years ago. This shows that dog numbers depended on human numbers.
Earlier, in 2018, a study suggested that the Yana site showed signs of wolves starting to be tamed. Remains of medium-sized dog-like animals were found there. They showed signs of living with people, like worn teeth and juvenile features in an almost-adult skull. These signs suggest that these animals were living with humans and were in the very early stages of domestication.
Mixing of Genes
Studies show that dog-wolf ancestors mixed with golden jackals. However, since domestication, there has been very little gene flow from wolves into dogs. But there has been a lot of gene flow from dogs into wolves. Some wolves were related to all ancient and modern dogs. A small amount of gene flow was found between coyotes and ancient American dogs, and between African wolves and African dogs.
The short time dogs and wolves separated, followed by their constant mixing, means that 20% of East Asian wolf DNA and 7–25% of European and Middle Eastern wolf DNA comes from dogs. The gene that causes black fur in North American wolves came from early Native American dogs. Dogs and wolves in the Himalayas and Tibetan plateau have a gene that helps them live in high altitudes. This gene came from an unknown wolf-like animal that is now extinct.
Bonn-Oberkassel Dog

The earliest widely accepted dog remains were found in Bonn-Oberkassel, Germany. Evidence from its context, bones, and DNA shows this dog was clearly not a local wolf. It was dated to 14,223 years ago.
In 1914, two human skeletons were found in Germany. With them was a jawbone of what was thought to be a "wolf." After World War I, in 1919, these remains were studied. The jawbone was called a wolf's and then stored away. In the late 1970s, scientists re-examined the jawbone and reclassified it as a domesticated dog. Its DNA matched Canis familiaris (a dog). The human bodies were dated to 14,223 years ago. This means that "modern" dogs, both in looks and genetics, existed in Western Europe around 14,500 years ago.
Later studies found more bones belonging to this dog, allowing scientists to put together most of its skeleton. The humans were a man (40 years old) and a woman (25 years old). All three remains were covered in red powder and large basalt blocks. It's believed that a dog was buried with the two humans. A tooth from a smaller, older dog was also found, but it wasn't covered in red powder.
The dog likely died young from canine distemper (a disease) between 19 and 23 weeks old. It couldn't have survived without a lot of human care. During this time, the dog wasn't useful for hunting or other tasks. This suggests that humans had strong emotional bonds with this dog. In short, near the end of the Ice Age, some humans cared deeply for their dogs, not just for their usefulness.
Ice Age Dogs
In 2020, studies of ancient dog DNA showed that all dogs came from an ancient, extinct wolf group. By the end of the last Ice Age (11,700 years ago), five different dog groups had formed. These groups are seen in dog samples from the Neolithic era Levant (7,000 years ago), Mesolithic era Karelia (10,900 years ago), Mesolithic era Baikal (7,000 years ago), ancient America (4,000 years ago), and the New Guinea singing dog (still alive today).
The world's ancient and modern dog populations can be divided into Arctic/Americas, East Asian, and West Eurasian groups.
- The Arctic/Americas group includes modern arctic breeds, a 9,500-year-old dog from Zhokhov Island, ancient American dogs, and dogs from Siberia.
- The East Asian group includes modern dogs from China, Vietnam, and the dingo and New Guinea singing dog.
- The West Eurasian group includes ancient dogs from the Near East and Europe, modern African dogs, and Bronze Age dogs from the Eurasian Steppe.
Ancient and modern European dogs are more closely related to arctic dogs than to Near Eastern dogs. This shows a big mixing event in Europe. Dogs from Karelia (about 10,000 years ago) had ancestry from both Arctic (70%) and Western Eurasian (30%) groups. This means early European dogs already had mixed ancestry. The 9,500-year-old Zhokhov dog from Siberia had no Western Eurasian ancestry. This means Western dog ancestry hadn't reached the Siberian Arctic by then. The earliest Neolithic European dog (7,000 years old) was a mix of Karelian and Levantine groups. A 5,000-year-old dog from Sweden was the ancestor of almost all modern European dogs. This suggests that a mixed group of dogs replaced all other dog populations in Europe.
Siberian dogs were genetically similar 9,500–7,000 years ago, showing Arctic ancestry. But dogs from the Eurasian Steppe and Europe later mixed with them. This led to Siberian dogs having different levels of Arctic and Near East ancestry. Dogs from the Bronze Age Eurasian Steppe had 40% ancient arctic and 60% ancient Near East ancestry until the Middle Ages. This suggests that dogs moved with the spread of farming from the Near East into the steppes.
Ancient human DNA and ancient dog DNA generally matched each other. This means that as people moved, their dogs often moved with them. But there were some differences. For example, the same dogs could be found in the Neolithic Levant and later in Iran, even though the human populations were different. This implies that dogs sometimes moved between human groups, perhaps as trade items. Dogs seem to have spread across Eurasia and into the Americas without major human population movements. This is still a mystery.
The AMY2B gene helps digest starch. At the start of farming, only some dogs had this gene. It became widespread several thousand years later.
Dogs traveled alongside humans, but their movements didn't always match. This means that sometimes humans moved without dogs, or dogs moved between human groups. Past studies suggested where dogs came from, but these studies were based on modern DNA patterns. The true origin of dogs is still unclear because of recent gene mixing and population changes.
First Dogs as a Hunting Tool

During the Upper Paleolithic (50,000–10,000 years ago), human populations grew, hunting tools improved, and the climate changed. This might have reduced prey animals. Scavenging became very important for some wolf groups to survive. Changes like tameness and smaller body size would make them less efficient hunters, leading them to rely on scavenging. We don't know if these early dogs were just scavengers living near humans, or if they were companions or hunters that helped them spread.
Scientists have suggested that a hunting partnership between humans and dogs was key to dog domestication. Ancient rock art from Saudi Arabia (8,000 years ago) shows many dogs hunting with humans. Some dogs are even on leashes. As the climate changed from cold and dry to warmer and wetter, forests grew. It's thought that hunting dogs were used to track and find wounded game in thick forests. Dogs' ability to chase, track, sniff, and hold prey greatly increases hunting success in forests, where human senses are not as sharp. Dogs are still used for hunting in forests today.
Arctic Breeds
First Dog Breeds in Arctic Northeastern Siberia

Domestic dogs were present 9,500 years ago on Zhokhov Island, in arctic northeastern Siberia. Discoveries at this site include dog harness straps, similar to those used by modern Inuit. There are also bones of polar bears and reindeer, suggesting long hunting trips and carrying large animal parts back to the site. Tools made from obsidian (a type of rock) were brought from 1,500 kilometers away. These findings suggest that sled dogs were used for long-distance transport.
A study of dog remains shows that these dogs were specially bred. Some were bred as sled dogs, and others as hunting dogs. This means that people had specific standards for these dog types back then. The best size for a sled dog is 16 to 25 kg, and the ancient sled dogs fit this. The same size standard is seen in sled dogs from 2,000 years ago and in the modern Siberian husky. Other dogs were larger (30 kg) and might have been crossed with wolves for polar bear hunting. When these dogs died, their heads were carefully removed from their bodies by humans, possibly for ceremonies.
The study suggests that dog evolution happened in three stages after they separated from wolves. First, natural selection based on feeding behavior near humans. Second, artificial selection based on tameness. Third, specific breeding for tasks like sled pulling. This process started 30,000–40,000 years ago and sped up until domestication was complete.
The Zhokhov dogs are the oldest known dogs to show different coat colors. They had black patterns on their backs, which helped tell them apart from white arctic wolves.
Dogs Enter North America from Northeastern Siberia
Evidence shows that dogs were harnessed in the Arctic 9,000 years ago. DNA from these dogs shows they are related to modern Arctic dogs. This lineage gave rise to the first native American dogs. Later, other types of dogs were brought by the Thule people and European settlers. European dogs replaced the older dog groups in North America.
In North America, the oldest dog remains were found in Alaska, dating to 10,150 years ago. DNA suggests this dog's lineage split from the Siberian Zhokhov Island dog lineage 16,700 years ago. This matches when the North Pacific coastal route to North America might have opened. Analysis of its bones suggests it ate a marine diet. The next oldest dogs were found in Illinois, dating to 9,900 years ago. These were medium-sized, active dogs that ate mostly freshwater fish. Similar dog burials in Eurasia suggest dogs were important for hunting. They gained a high social status.
A 2018 study found that North American dog fossils were most closely related to a 9,000-year-old fossil from Zhokhov Island, Siberia. The study suggested that all North American dogs shared a common ancestor from 14,600 years ago. This ancestor, along with the Zhokhov dog's ancestor, split from a common ancestor 15,600 years ago. The Koster dogs show that dogs entered North America from Siberia 4,500 years after humans. They were isolated for 9,000 years. After Europeans arrived, these original American dogs disappeared, replaced by Eurasian dogs. However, their unique genetic signature lives on in arctic breeds like Alaskan malamutes and Siberian huskies.
In 2019, a study found that the first dogs brought to the North American Arctic were later replaced by dogs that came with the Inuit 2,000 years ago. These Inuit dogs were more diverse. Today, Arctic sled dogs are some of the last descendants of these pre-European dog lineages in the Americas. In 2020, DNA sequencing showed that two Mexican breeds, the Chihuahua and the Xoloitzcuintli, still have a small amount of this pre-colonial ancestry.
Late Pleistocene Wolf Mixing

In 2015, scientists mapped the DNA of a 35,000-year-old Ice Age wolf fossil from the Taymyr Peninsula in arctic Siberia. They compared it to modern dogs and grey wolves. This Taimyr wolf was equally related to both dogs and modern wolves. But it shared more genes with high-latitude and arctic dog breeds like the Siberian husky and Greenland dog. The Greenland dog has 3.5% Taimyr wolf ancestry. This means there was mixing between the Taimyr wolf population and the ancestors of these four high-latitude dog breeds. This mixing might have helped early dogs in cold places adapt to the harsh environment. It also suggests that modern dog breeds came from more than one region.
Since the Taimyr wolf contributed to Arctic breeds, it means that descendants of the Taimyr wolf survived until dogs were tamed in Europe. When dogs arrived in high latitudes, they mixed with local wolves. Both contributed to modern Arctic breeds. Based on the oldest dog remains, domestic dogs likely arrived in high latitudes within the last 15,000 years.
The DNA of a dog found in Ireland, dated 4,800 years ago, was also studied. This dog was male and didn't have genes for modern coat length or color. It could process starch better than wolves, but not as well as modern dogs. It also had ancestry from a wolf population not found in other dogs or wolves today. The DNA suggests that modern wolf and dog populations separated between 20,000 and 60,000 years ago. This means dogs might have been tamed much earlier than we thought, or they arrived in the Arctic early, or both. Another idea is that because northern breeds have some Taimyr wolf ancestry, there might have been more than one domestication event.
In 2020, the DNA of a 33,000-year-old Ice Age wolf from the Yana River in arctic Siberia was studied. This Yana wolf was more closely related to the 35,000-year-old Taimyr wolf than to modern wolves. There was evidence of gene flow between the Yana-Taimyr wolves and early dogs like the Pre-Columbian, Zhokhov, and modern sled dogs. This suggests that ancient wolves mixed their genes with the ancestors of these dogs. There has been no evidence of mixing between sled dogs and modern grey wolves for the past 9,500 years. Greenland sled dogs have been kept separate from other breeds for 850 years. Their DNA shows more history with the Zhokhov dogs than any other arctic breed. Sled dogs don't show an adaptation to a starch-rich diet like other dogs. But they do show an adaptation to a high intake of fat and fatty acids, which wasn't found in the Zhokhov dogs. This suggests that sled dogs adapted to the low-starch, high-fat diet of the people they lived with.
In 2021, a study of four more ancient Siberian wolf DNA sequences showed they were similar to the Taimyr and Yana wolves. These six extinct wolves branched off from the lineage that leads to modern wolves and dogs. All six ancient wolves shared genes with Arctic dogs: Greenland dogs, Siberian and Alaskan huskies, Alaskan malamutes, the extinct Zhokhov dog, and the extinct pre-European contact dogs of North America. It's possible that another extinct wolf population, related to all six, contributed to the ancestry of Arctic dogs.
Dogs Enter Japan
The oldest dog fossil found in Japan is 9,500 years old. As the weather got warmer after the Ice Age, forests grew in Japan. People changed from hunting large animals to hunting faster sika deer and wild boar in dense forests. This led to changes in hunting tools, like smaller arrowheads. A study of the Jōmon people who lived in Japan during this time shows they buried dogs individually. They probably used dogs to hunt deer and wild boar, just as hunters in Japan still do today.
Hunting dogs are very important to hunter-gatherer societies. Records show they were given names, treated as family, and seen as different from other dogs. They were often buried separately with markers and grave goods. Exceptional hunters or dogs killed during a hunt were often honored. A dog's value as a hunting partner gave them high status, like a living weapon. The most skilled dogs were seen as having "personhood," with a social position similar to skilled human hunters.
Intentional dog burials with hunting evidence are also found in early forest hunter-gatherer societies in Europe and North America. This shows that hunting dogs were widely used in temperate forest areas across the Northern Hemisphere.
Dogs Enter Island Southeast Asia and Oceania
.png)
.jpg)
In 2020, a DNA study of ancient dog fossils from southern China showed that most belonged to a group called haplogroup A1b. This group is also found in Australian dingoes and pre-colonial Pacific dogs, but it's rare in China today. The oldest sample from this group, from China, dates to 7,000 years ago. Dogs from this group were once common in southern China. They then spread through Southeast Asia into New Guinea and Oceania. But they were replaced in China 2,000 years ago by dogs from other groups.
Dogs entered Island Southeast Asia, Australia, and New Guinea at least twice. One group arrived with ancient sea-faring hunter-gatherers between 10,000 and 5,000 years ago. Another group arrived with later farming and trading people called Austronesian peoples (from Taiwan) around 5,000 years ago. The dogs that came with farmers could digest starch. This shows they traveled with humans who grew cereal crops. The oldest archaeological remains of dogs in Island Southeast Asia and Oceania are a dog burial in Timor and dingo remains in Australia, both about 3,500 years old. The Timor dog is thought to be from the second wave, and the dingo from the first. Austronesian dogs were highly valued as hunting companions, especially for wild boar. In the Philippines, dog remains have been found buried near human graves.
From Island Southeast Asia, Austronesian sailors carried dogs into nearby islands. However, on smaller islands in Island Melanesia and Polynesia, dogs lost their importance as hunting animals. This is because there were no wild boars or other large mammals to hunt. Dogs then became competitors for limited food and were themselves eaten. Many of the original Austronesian dogs were eaten to extinction on these islands.
This caused a break in the genes of tamed dogs and the words for "dog" among Austronesians in the Pacific Islands. However, dogs were later brought back from neighboring groups and carried further east into Polynesia, reaching as far as Hawaii. Even though these dogs were used as food, not hunting companions. Genetic studies confirm this, showing that Polynesian dogs came from the first wave of dog introductions and are not related to dogs from Taiwan and the Philippines.
Dogs from the Near East Enter Africa
In 2020, ancient dog DNA showed that modern dogs in sub-Sahara Africa came from a single origin in the Levant. An ancient dog from there was dated to 7,000 years ago. This matches how humans moved from the Levant into Africa during the Neolithic period, bringing cattle with them. Since then, there has been little gene flow into African dogs until the last few hundred years. Descendants of a dog from Iran (5,800 years ago) and dogs from Europe completely replaced the Levant dog lineage 2,300 years ago. This was linked to human migration from Iran and some smaller migrations from Europe. Today, all Near Eastern dogs show 81% ancient Iranian and 19% Neolithic European ancestry.
The oldest dog remains found in Africa are 5,900 years old, from Egypt. The next oldest are 5,500 years old, from Sudan. This suggests that dogs arrived from Asia at the same time as domestic sheep and goats. Dogs then spread south through Africa with livestock herders. In 2020, ancient dog DNA showed that the southern African Rhodesian Ridgeback still has 4% pre-colonial ancestry.
Images for kids
-

The dog diverged from a now-extinct population of wolves 27,000–40,000 years ago immediately before the Last Glacial Maximum, when much of the mammoth steppe was cold and dry.
-

Cave wolf skull, Natural History Museum, Berlin
-
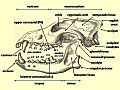
Diagram of a wolf skull with key features labelled
-

Mandible of Canis c.f. variabilis from northeastern Siberia dated 360,000–400,000 years old
-

Skull of the "Altai dog" that is dated 33,300 years old
-

The 14,500-year-old upper-right jaw of a Pleistocene wolf found in the Kessleroch cave near Thayngen in the canton of Schaffhausen, Switzerland
-

Watercolor tracing made by archaeologist Henri Breuil from a cave painting of a wolf-like canid, Font-de-Gaume, France, dated 19,000 years ago
-

Reduction in size under selective breeding – grey wolf and chihuahua skulls
-

The difference in overall body size between a Cane Corso (Italian mastiff) and a Yorkshire terrier is over 30-fold, yet both are members of the same species.
-

Mammoth bone dwelling, Mezhirich, Cherkasy Oblast, Ukraine
-

Bison surrounded by grey wolf pack
-

Example of a dog making prolonged eye contact with a human
-

Mandible of the oldest recognised dog discovered in Oberkassel, Bonn, Germany, and dated 14,200 years old
-
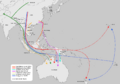
Map depicting possible phylogeographic origins of dingoes, New Guinea singing dogs, and Island Southeast Asian and Oceanian dogs, based on recent genetic evidence (Fillios & Taçon, 2016)
-

Ancient Hawaiian petroglyphic depiction of an ʻīlio (Hawaiian Poi Dog)
-

Sled dog types, sketched in 1833
-

The Greenland dog carries 3.5% genetic material inherited from a 35,000-year-old wolf from the Taymyr Peninsula, Arctic Siberia.
-

Petroglyph depicting two dogs hunting – Tassili n'Ajjer, Algeria
_Wolf.png)

